

Deconstructing gastrulation at single-cell resolution
- PMID: 35290798
- PMCID: PMC9221752
- DOI: 10.1016/j.cub.2022.02.059
Deconstructing gastrulation at single-cell resolution
Abstract
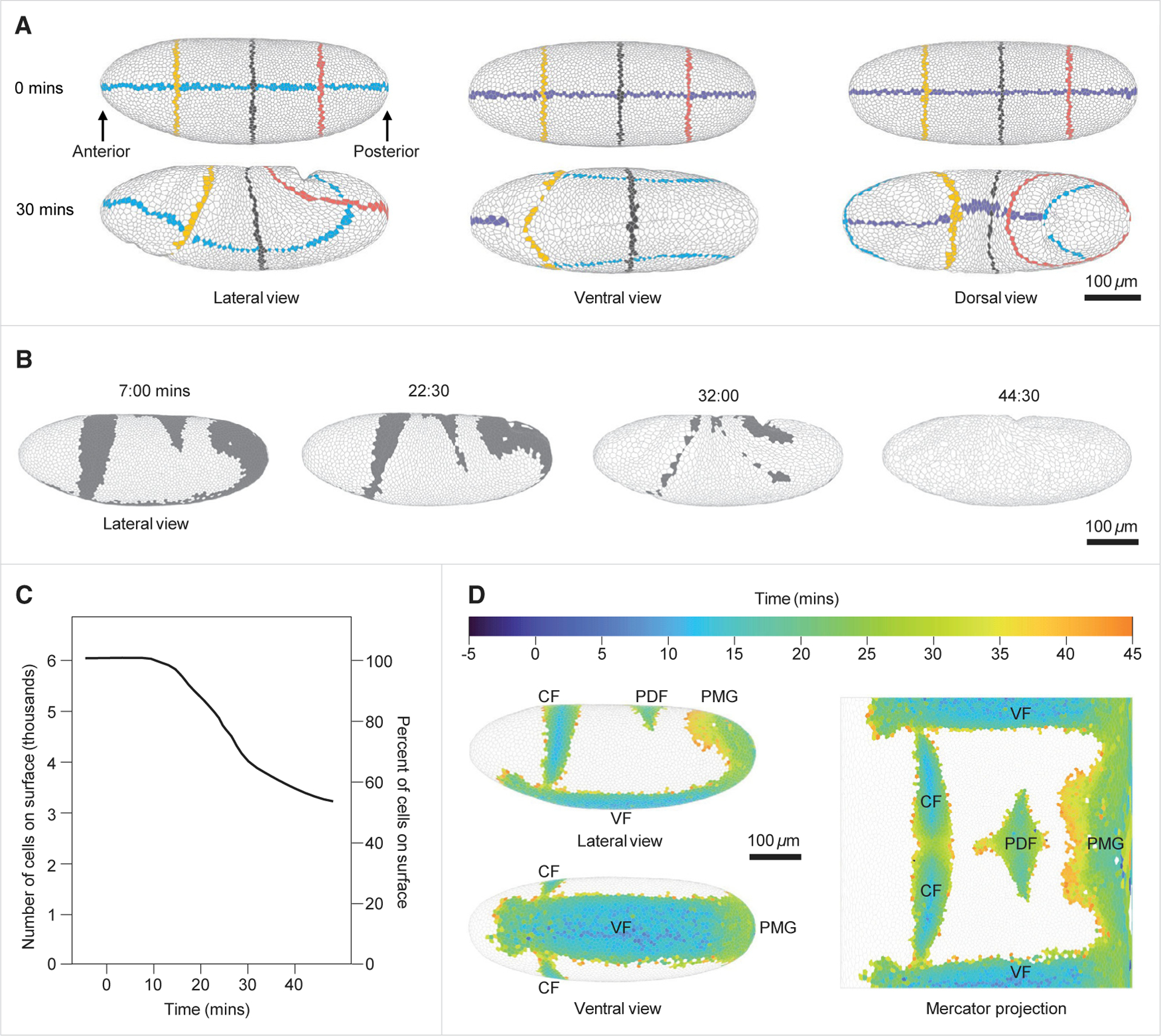
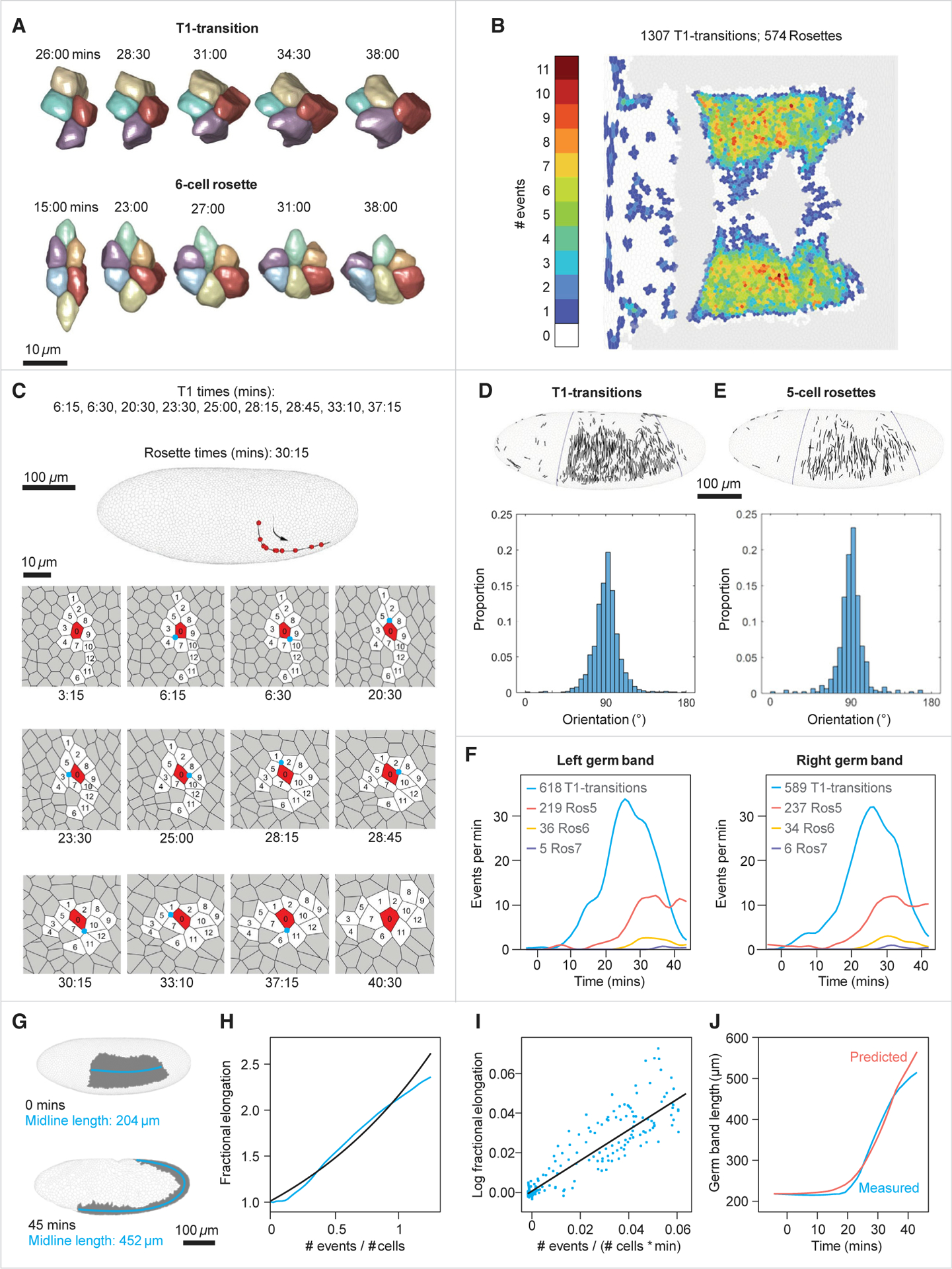
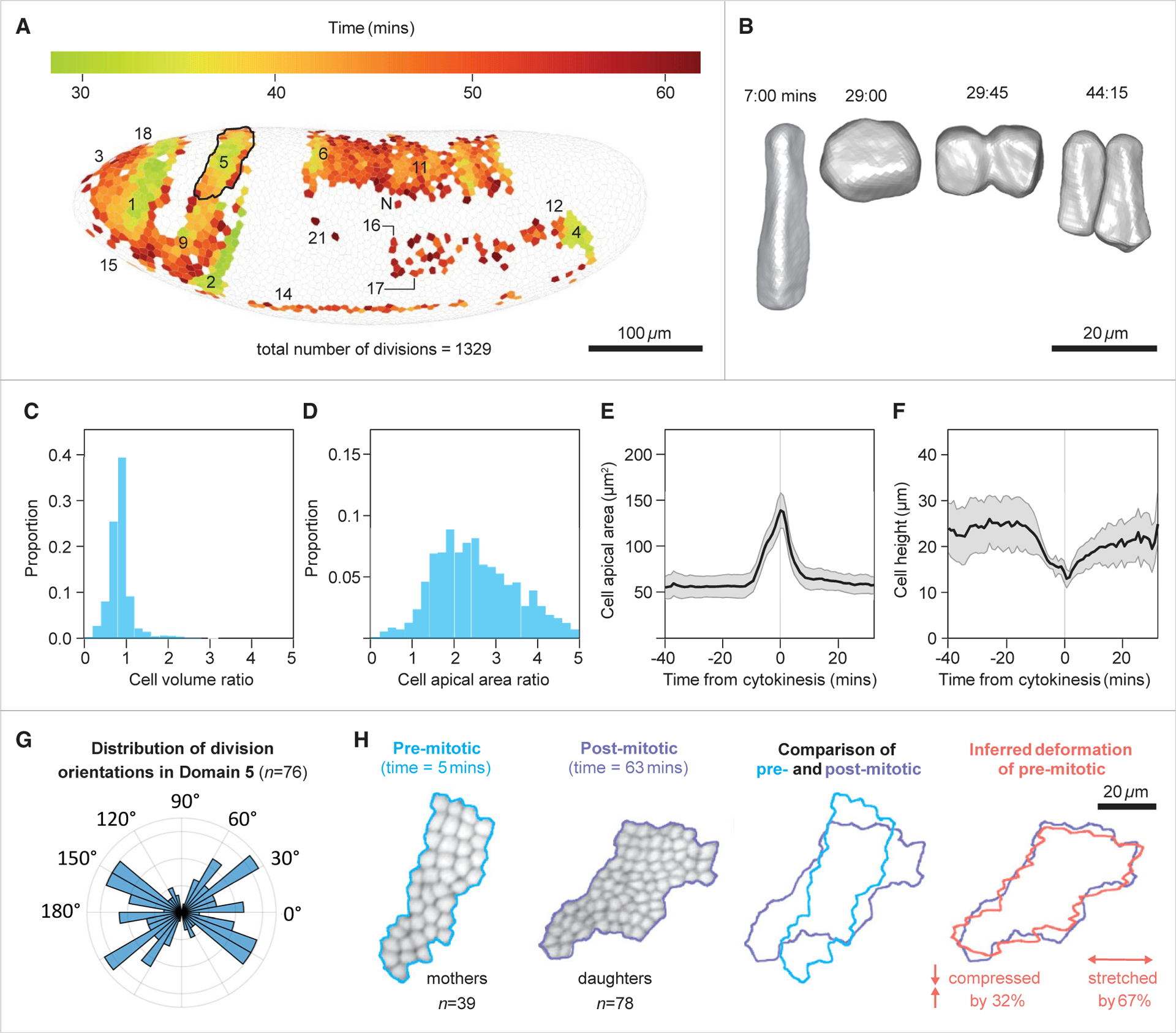
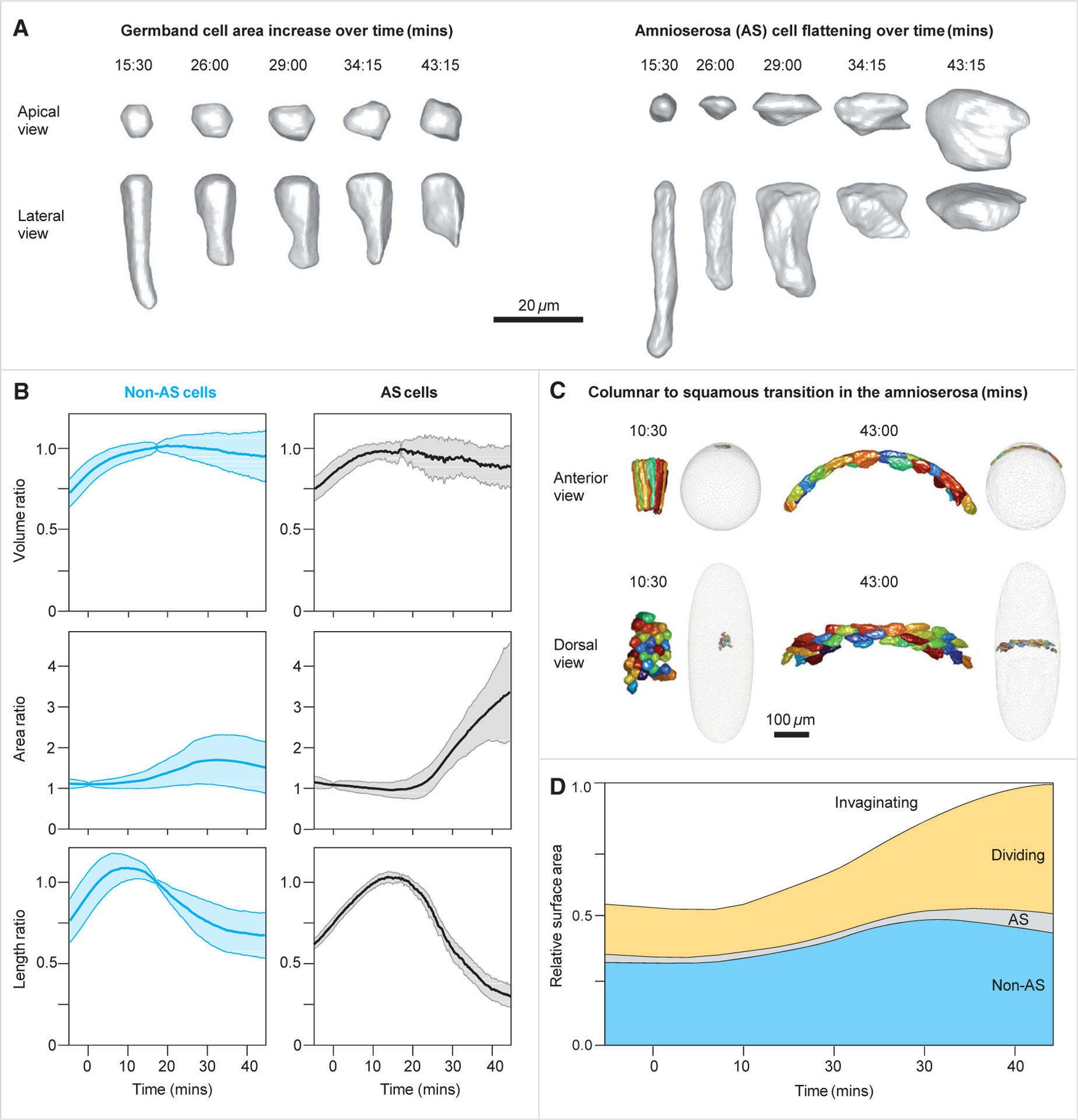
Gastrulation movements in all animal embryos start with regulated deformations of patterned epithelial sheets, which are driven by cell divisions, cell shape changes, and cell intercalations. Each of these behaviors has been associated with distinct aspects of gastrulation1-4 and has been a subject of intense research using genetic, cell biological, and more recently, biophysical approaches.5-14 Most of these studies, however, focus either on cellular processes driving gastrulation or on large-scale tissue deformations.15-23 Recent advances in microscopy and image processing create a unique opportunity for integrating these complementary viewpoints.24-28 Here, we take a step toward bridging these complementary strategies and deconstruct the early stages of gastrulation in the entire Drosophila embryo. Our approach relies on an integrated computational framework for cell segmentation and tracking and on efficient algorithms for event detection. The detected events are then mapped back onto the blastoderm shell, providing an intuitive visual means to examine complex cellular activity patterns within the context of their initial anatomic domains. By analyzing these maps, we identified that the loss of nearly half of surface cells to invaginations is compensated primarily by transient mitotic rounding. In addition, by analyzing mapped cell intercalation events, we derived direct quantitative relations between intercalation frequency and the rate of axis elongation. This work is setting the stage for systems-level dissection of a pivotal step in animal development.
Copyright © 2022. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Irvine KD, and Wieschaus E (1994). Cell intercalation during Drosophila germband extension and its regulation by pair-rule segmentation genes. Development 120, 827–841. - PubMed
-
- Foe VE (1989). Mitotic domains reveal early commitment of cells in Drosophila embryos. Trends Genet. 5, 322. - PubMed
-
- Pope KL, and Harris TJC (2008). Control of cell flattening and junctional remodelling during squamous epithelial morphogenesis in Drosophila. Development 135, 2227–2238. - PubMed
-
- Sweeton D, Parks S, Costa M, and Wieschaus E (1991). Gastrulation in Drosophila: the formation of the ventral furrow and posterior midgut invaginations. Development 112, 775–789. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
