

Human and Insect Cell-Produced Recombinant Adeno-Associated Viruses Show Differences in Genome Heterogeneity
- PMID: 35293222
- PMCID: PMC9063199
- DOI: 10.1089/hum.2022.050
Human and Insect Cell-Produced Recombinant Adeno-Associated Viruses Show Differences in Genome Heterogeneity
Abstract
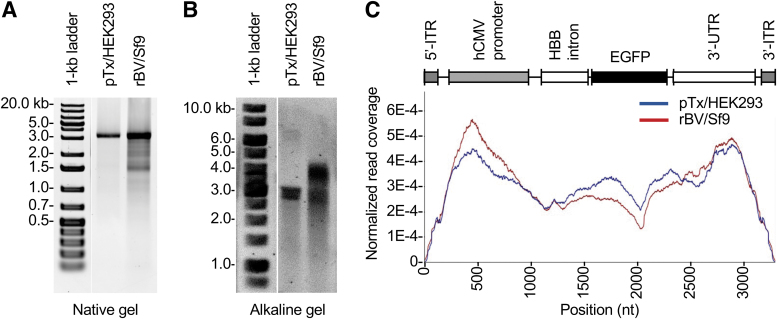
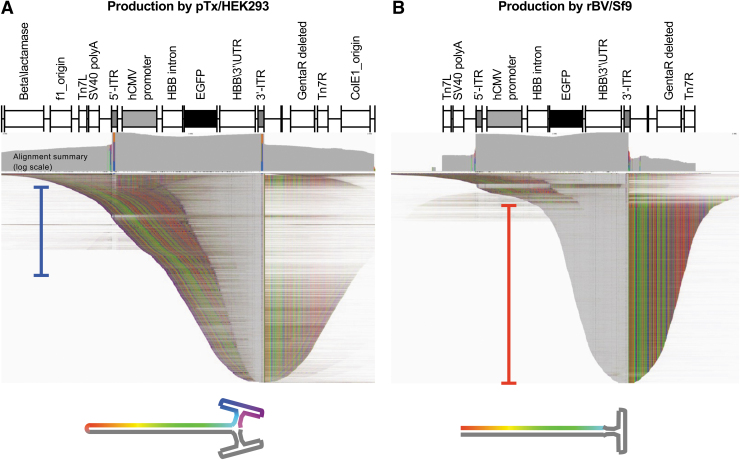
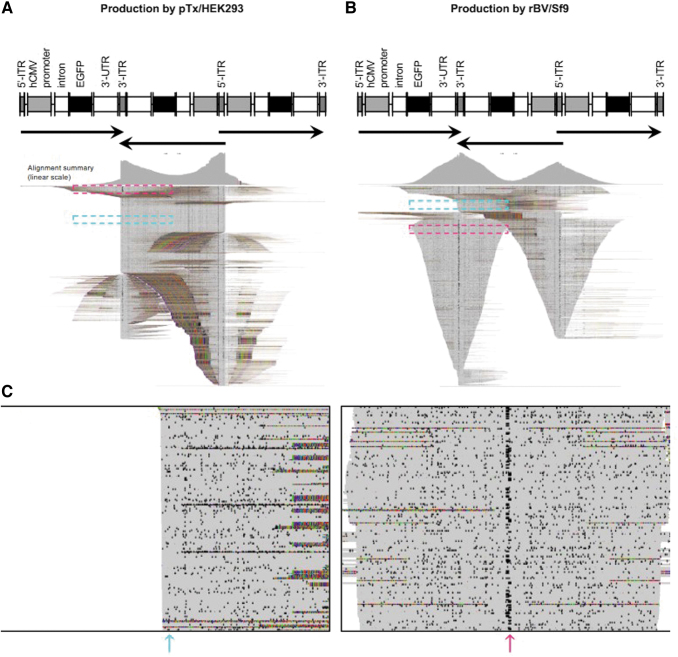
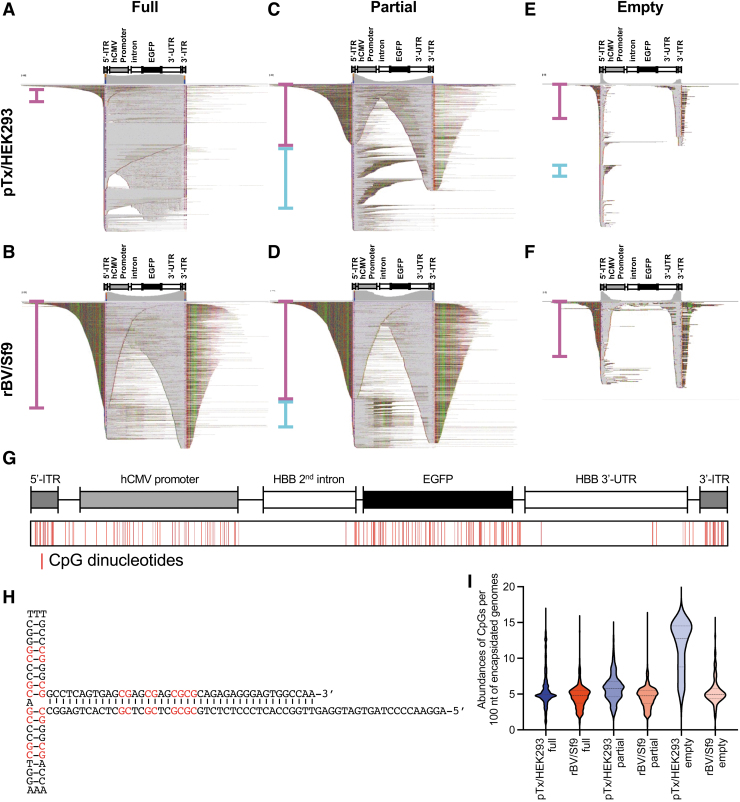
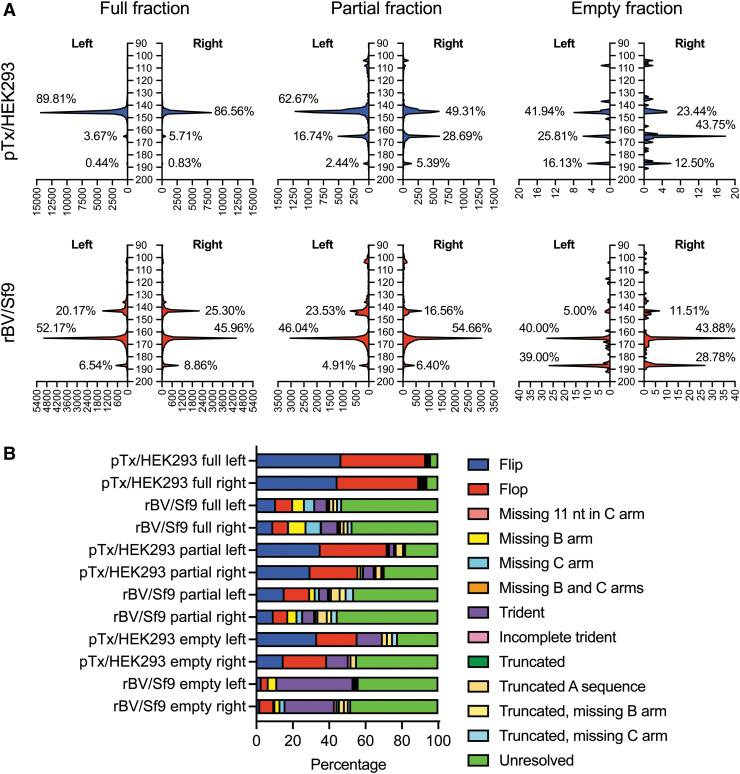
In the past two decades, adeno-associated virus (AAV) vector manufacturing has made remarkable advancements to meet large-scale production demands for preclinical and clinical trials. In addition, AAV vectors have been extensively studied for their safety and efficacy. In particular, the presence of empty AAV capsids and particles containing "inaccurate" vector genomes in preparations has been a subject of concern. Several methods exist to separate empty capsids from full particles; but thus far, no single technique can produce vectors that are free of empty or partial (non-unit length) capsids. Unfortunately, the exact genome compositions of full, intermediate, and empty capsids remain largely unknown. In this work, we used AAV-genome population sequencing to explore the compositions of DNase-resistant, encapsidated vector genomes produced by two common production pipelines: plasmid transfection in human embryonic kidney cells (pTx/HEK293) and baculovirus expression vectors in Spodoptera frugiperda insect cells (rBV/Sf9). Intriguingly, our results show that vectors originating from the same construct design that were manufactured by the rBV/Sf9 system produced a higher degree of truncated and unresolved species than those generated by pTx/HEK293 production. We also demonstrate that empty particles purified by cesium chloride gradient ultracentrifugation are not truly empty but are instead packaged with genomes composed of a single truncated and/or unresolved inverted terminal repeat (ITR). Our data suggest that the frequency of these "mutated" ITRs correlates with the abundance of inaccurate genomes in all fractions. These surprising findings shed new light on vector efficacy, safety, and how clinical vectors should be quantified and evaluated.
Keywords: HEK293; Sf9; adeno-associated virus; gene therapy; vector heterogeneity.
Conflict of interest statement
G.G. is a scientific cofounder of Voyager Therapeutics and Aspa Therapeutics and holds equity in these companies. G.G. is an inventor on patents with potential royalties licensed to Voyager Therapeutics, Aspa Therapeutics, and other biopharmaceutical companies. The remaining authors declare no competing interests. M.P.-B. and E.A. are inventors of patents related to AAV gene therapy licensed to biopharma companies. K.W. is a full-time employee of Pacific Biosciences, a company commercializing SMRT sequencing technologies.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
