

The Signature of Moderate Perinatal Hypoxia on Cortical Organization and Behavior: Altered PNN-Parvalbumin Interneuron Connectivity of the Cingulate Circuitries
- PMID: 35295859
- PMCID: PMC8919082
- DOI: 10.3389/fcell.2022.810980
The Signature of Moderate Perinatal Hypoxia on Cortical Organization and Behavior: Altered PNN-Parvalbumin Interneuron Connectivity of the Cingulate Circuitries
Abstract
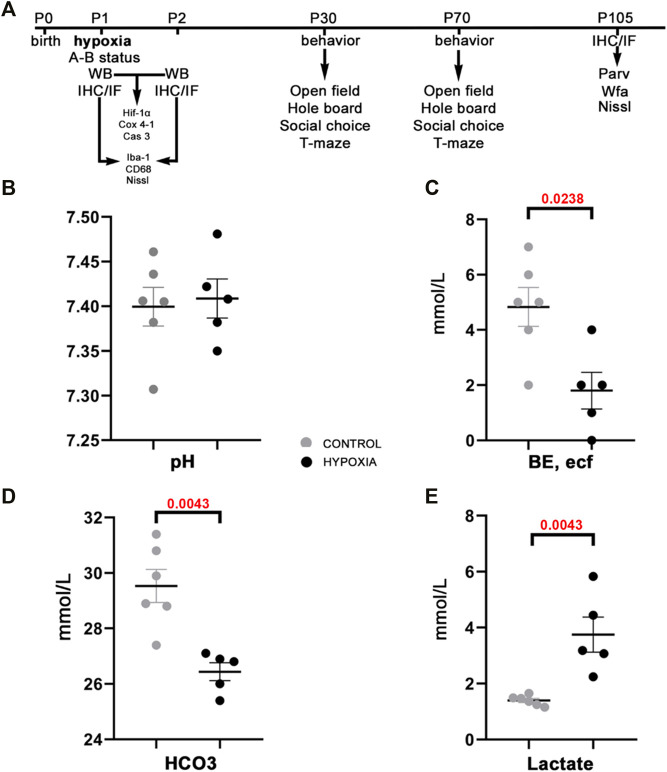
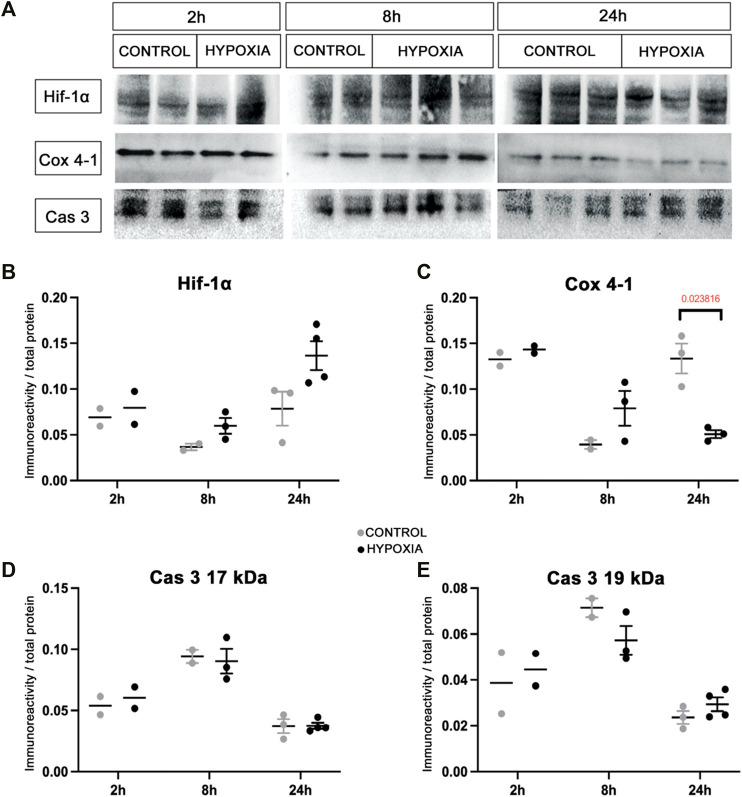
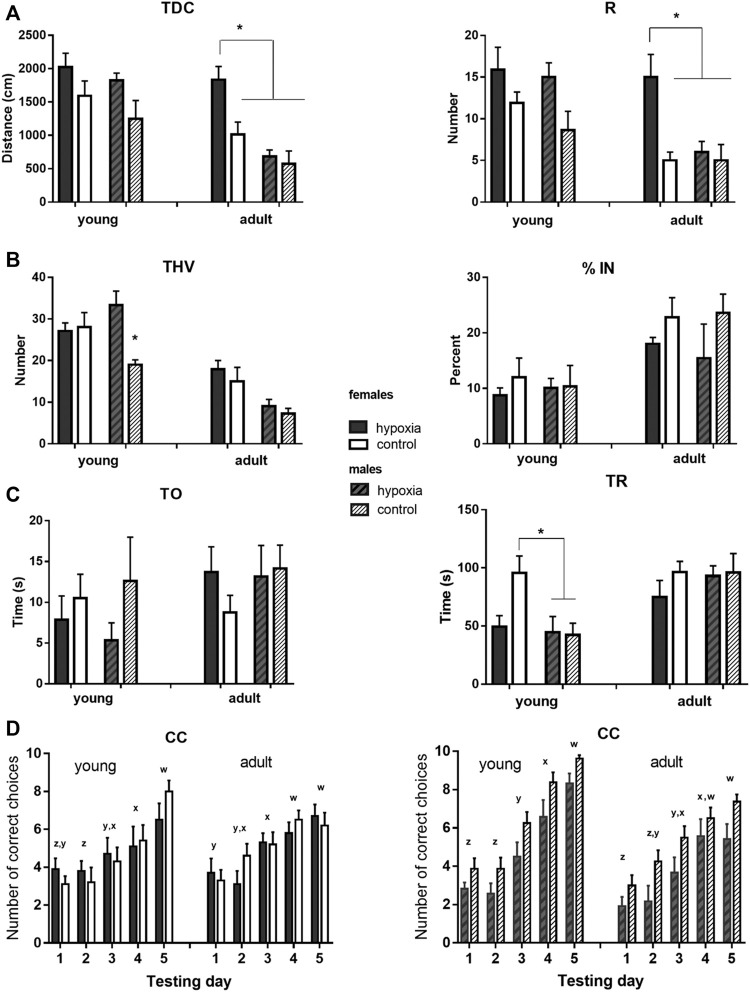
This study was designed in a rat model to determine the hallmarks of possible permanent behavioral and structural brain alterations after a single moderate hypoxic insult. Eighty-two Wistar Han (RccHan: WIST) rats were randomly subjected to hypoxia (pO2 73 mmHg/2 h) or normoxia at the first postnatal day. The substantially increased blood lactate, a significantly decreased cytochrome-C-oxygenase expression in the brain, and depleted subventricular zone suggested a high vulnerability of subset of cell populations to oxidative stress and consequent tissue response even after a single, moderate, hypoxic event. The results of behavioral tests (open-field, hole-board, social-choice, and T-maze) applied at the 30-45th and 70-85th postnatal days revealed significant hyperactivity and a slower pace of learning in rats subjected to perinatal hypoxia. At 3.5 months after hypoxic insult, the histochemical examination demonstrated a significantly increased number of specific extracellular matrix-perineuronal nets and increased parvalbumin expression in a subpopulation of interneurons in the medial and retrosplenial cingulate cortex of these animals. Conclusively, moderate perinatal hypoxia in rats causes a long-lasting reorganization of the connectivity in the cingulate cortex and consequent alterations of related behavioral and cognitive abilities. This non-invasive hypoxia model in the rat successfully and complementarily models the moderate perinatal hypoxic injury in fetuses and prematurely born human babies and may enhance future research into new diagnostic and therapeutic strategies for perinatal medicine.
Keywords: cortical development; hyperactivity behaviors; learning disabilities; oxidative stress; plasticity.
Copyright © 2022 Trnski, Nikolić, Ilic, Drlje, Bobic-Rasonja, Darmopil, Petanjek, Hranilovic and Jovanov-Milosevic.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
A lack of GluN2A-containing NMDA receptors confers a vulnerability to redox dysregulation: Consequences on parvalbumin interneurons, and their perineuronal nets.Neurobiol Dis. 2018 Jan;109(Pt A):64-75. doi: 10.1016/j.nbd.2017.10.006. Epub 2017 Oct 10. Neurobiol Dis. 2018. PMID: 29024713
-
Parvalbumin Interneurons and Perineuronal Nets in the Hippocampus and Retrosplenial Cortex of Adult Male Mice After Early Social Isolation Stress and Perinatal NMDA Receptor Antagonist Treatment.Front Synaptic Neurosci. 2021 Sep 22;13:733989. doi: 10.3389/fnsyn.2021.733989. eCollection 2021. Front Synaptic Neurosci. 2021. PMID: 34630066 Free PMC article.
-
Chronic Stress Modulates Interneuronal Plasticity: Effects on PSA-NCAM and Perineuronal Nets in Cortical and Extracortical Regions.Front Cell Neurosci. 2019 May 7;13:197. doi: 10.3389/fncel.2019.00197. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31133813 Free PMC article.
-
Calcium-binding proteins in the human developing brain.Adv Anat Embryol Cell Biol. 2002;165:III-IX, 1-92. Adv Anat Embryol Cell Biol. 2002. PMID: 12236093 Review.
-
The Chemorepulsive Protein Semaphorin 3A and Perineuronal Net-Mediated Plasticity.Neural Plast. 2016;2016:3679545. doi: 10.1155/2016/3679545. Epub 2016 Jan 14. Neural Plast. 2016. PMID: 27057361 Free PMC article. Review.
Cited by
-
Morphological and Molecular Characteristics of Perineuronal Nets in the Human Prefrontal Cortex-A Possible Link to Microcircuitry Specialization.Mol Neurobiol. 2025 Jan;62(1):1094-1111. doi: 10.1007/s12035-024-04306-1. Epub 2024 Jul 3. Mol Neurobiol. 2025. PMID: 38958887 Free PMC article.
-
Lasting mesothalamic dopamine imbalance and altered exploratory behavior in rats after a mild neonatal hypoxic event.Front Integr Neurosci. 2024 Jan 17;17:1304338. doi: 10.3389/fnint.2023.1304338. eCollection 2023. Front Integr Neurosci. 2024. PMID: 38304737 Free PMC article.
-
Extracellular Matrix Regulation in Physiology and in Brain Disease.Int J Mol Sci. 2023 Apr 11;24(8):7049. doi: 10.3390/ijms24087049. Int J Mol Sci. 2023. PMID: 37108212 Free PMC article. Review.
-
Adolescent-like Processing of Behaviorally Salient Cues in Sensory and Prefrontal Cortices of Adult Preterm-Born Mice.Res Sq [Preprint]. 2024 Dec 11:rs.3.rs-5529783. doi: 10.21203/rs.3.rs-5529783/v1. Res Sq. 2024. PMID: 39711564 Free PMC article. Preprint.
-
Cortical interneurons in schizophrenia - cause or effect?Croat Med J. 2023 Apr 30;64(2):110-122. doi: 10.3325/cmj.2023.64.110. Croat Med J. 2023. PMID: 37131313 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources