

Trinucleotide CGG Repeat Diseases: An Expanding Field of Polyglycine Proteins?
- PMID: 35295941
- PMCID: PMC8918734
- DOI: 10.3389/fgene.2022.843014
Trinucleotide CGG Repeat Diseases: An Expanding Field of Polyglycine Proteins?
Abstract
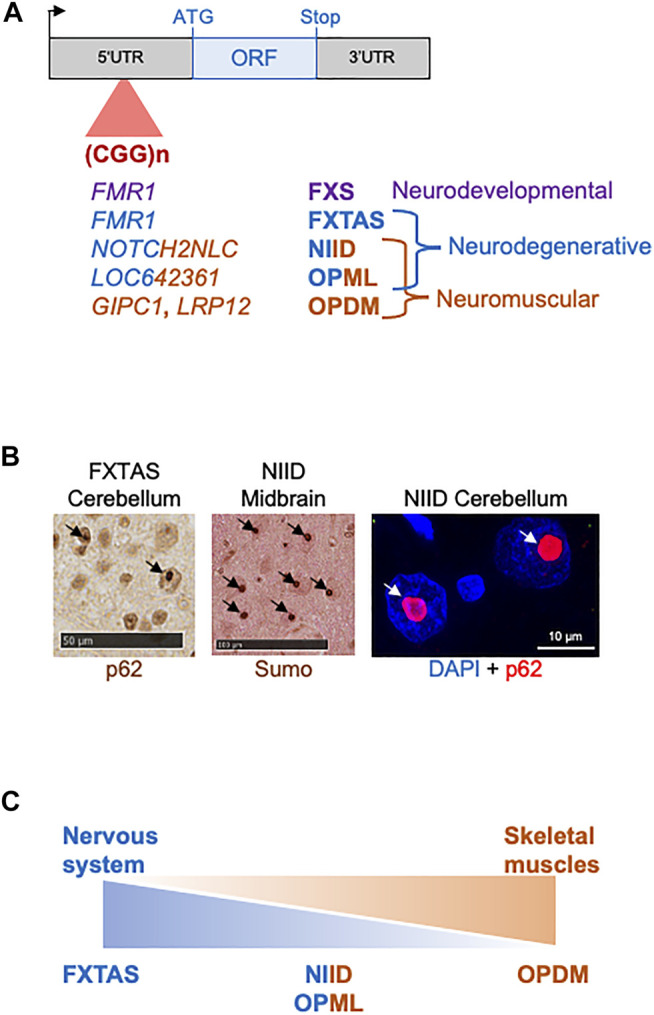
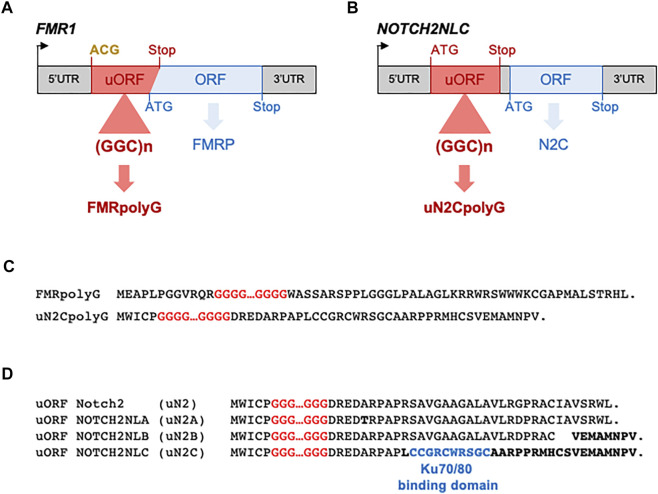
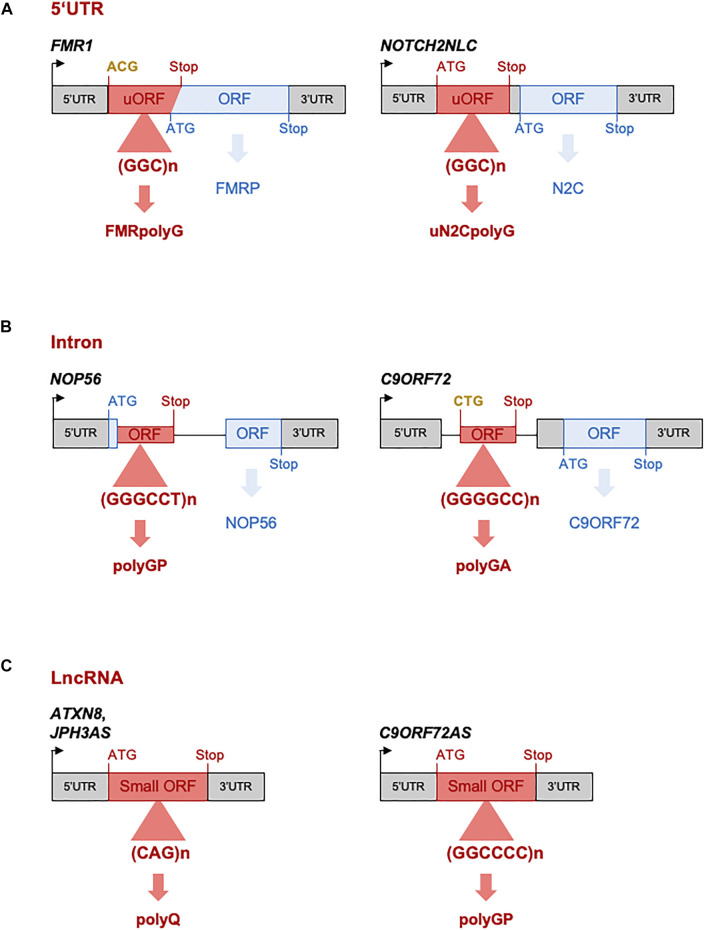
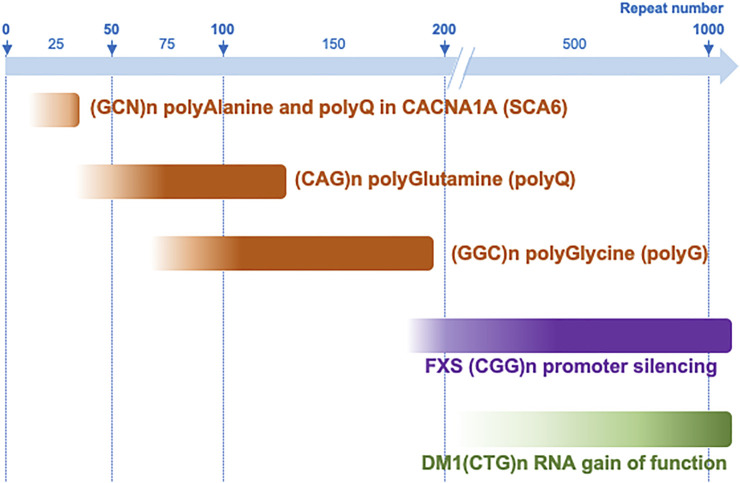
Microsatellites are repeated DNA sequences of 3-6 nucleotides highly variable in length and sequence and that have important roles in genomes regulation and evolution. However, expansion of a subset of these microsatellites over a threshold size is responsible of more than 50 human genetic diseases. Interestingly, some of these disorders are caused by expansions of similar sequences, sizes and localizations and present striking similarities in clinical manifestations and histopathological features, which suggest a common mechanism of disease. Notably, five identical CGG repeat expansions, but located in different genes, are the causes of fragile X-associated tremor/ataxia syndrome (FXTAS), neuronal intranuclear inclusion disease (NIID), oculopharyngodistal myopathy type 1 to 3 (OPDM1-3) and oculopharyngeal myopathy with leukoencephalopathy (OPML), which are neuromuscular and neurodegenerative syndromes with overlapping symptoms and similar histopathological features, notably the presence of characteristic eosinophilic ubiquitin-positive intranuclear inclusions. In this review we summarize recent finding in neuronal intranuclear inclusion disease and FXTAS, where the causing CGG expansions were found to be embedded within small upstream ORFs (uORFs), resulting in their translation into novel proteins containing a stretch of polyglycine (polyG). Importantly, expression of these polyG proteins is toxic in animal models and is sufficient to reproduce the formation of ubiquitin-positive intranuclear inclusions. These data suggest the existence of a novel class of human genetic pathology, the polyG diseases, and question whether a similar mechanism may exist in other diseases, notably in OPDM and OPML.
Keywords: RAN translation; microsatellite; neurodegeneration; protein aggregates toxicity; trinucleotide repeat disorders.
Copyright © 2022 Boivin and Charlet-Berguerand.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous