

Regulation of PGC1α Downstream of the Insulin Signaling Pathway Plays a Role in the Hepatic Proteotoxicity of Mutant α1-Antitrypsin Deficiency Variant Z
- PMID: 35301011
- PMCID: PMC9232923
- DOI: 10.1053/j.gastro.2022.03.010
Regulation of PGC1α Downstream of the Insulin Signaling Pathway Plays a Role in the Hepatic Proteotoxicity of Mutant α1-Antitrypsin Deficiency Variant Z
Abstract
Background & aims: Insulin signaling is known to regulate essential proteostasis mechanisms.
Methods: The analyses here examined effects of insulin signaling in the PiZ mouse model of α1-antitrypsin deficiency in which hepatocellular accumulation and proteotoxicity of the misfolded α1-antitrypsin Z variant (ATZ) causes liver fibrosis and cancer.
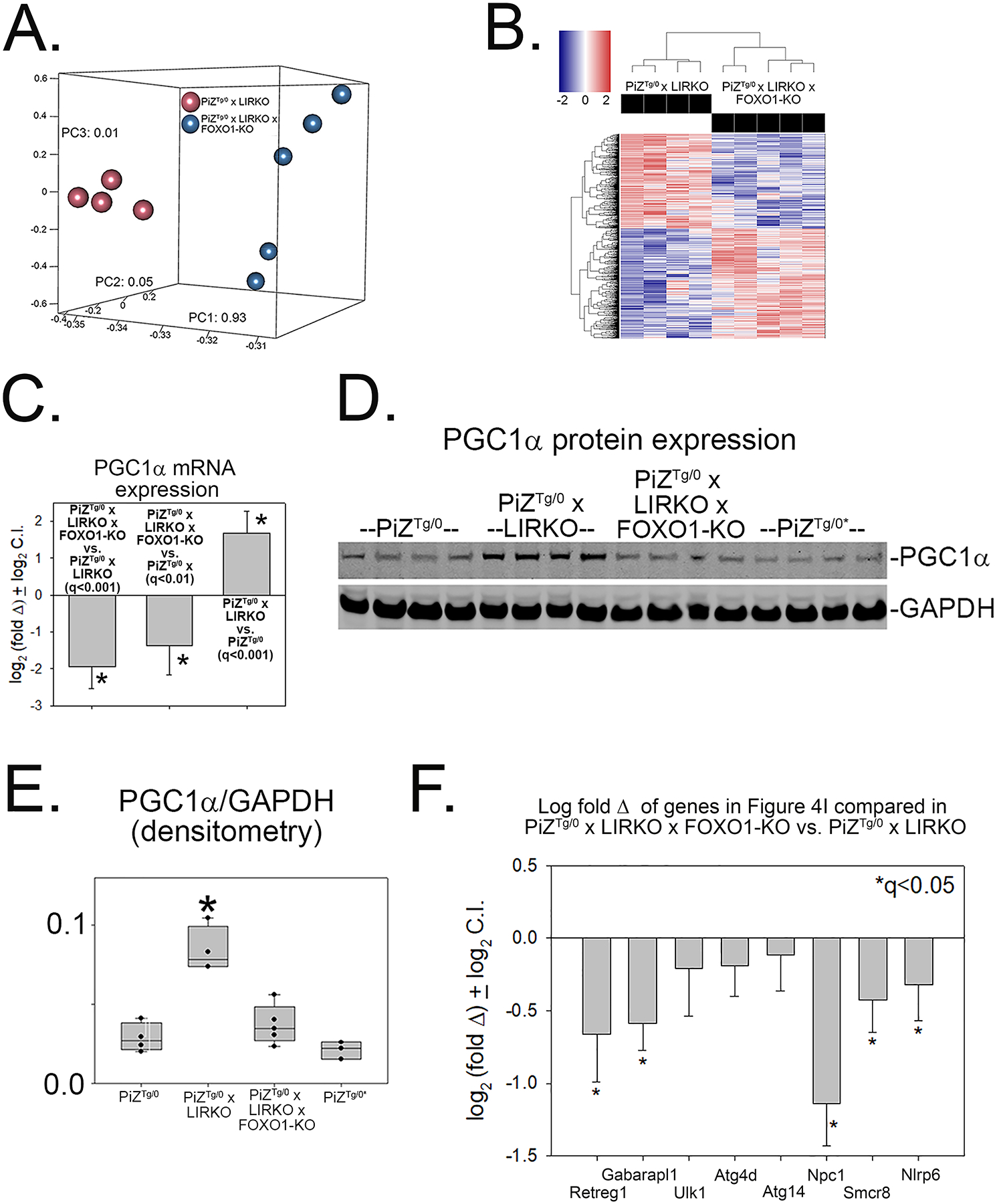
Results: We first studied the effects of breeding PiZ mice to liver-insulin-receptor knockout (LIRKO) mice (with hepatocyte-specific insulin-receptor gene disruption). The results showed decreased hepatic ATZ accumulation and liver fibrosis in PiZ x LIRKO vs PiZ mice, with reversal of those effects when we bred PiZ x LIRKO mice onto a FOXO1-deficient background. Increased intracellular degradation of ATZ mediated by autophagy was identified as the likely mechanism for diminished hepatic proteotoxicity in PiZ x LIRKO mice and the converse was responsible for enhanced toxicity in PiZ x LIRKO x FOXO1-KO animals. Transcriptomic studies showed major effects on oxidative phosphorylation and autophagy genes, and significant induction of peroxisome proliferator-activated-receptor-γ-coactivator-1α (PGC1α) expression in PiZ-LIRKO mice. Because PGC1α plays a key role in oxidative phosphorylation, we further investigated its effects on ATZ proteostasis in our ATZ-expressing mammalian cell model. The results showed PGC1α overexpression or activation enhances autophagic ATZ degradation.
Conclusions: These data implicate suppression of autophagic ATZ degradation by down-regulation of PGC1α as one mechanism by which insulin signaling exacerbates hepatic proteotoxicity in PiZ mice, and identify PGC1α as a novel target for development of new human α1-antitrypsin deficiency liver disease therapies.
Keywords: Autophagy; Liver Disease; Liver Fibrosis; Oxidative Phosphorylation; Proteostasis.
Copyright © 2022 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures





Comment in
-
A Role for Hepatic Insulin Signaling in α1-Antitrypsin Deficiency.Gastroenterology. 2022 Jul;163(1):49-51. doi: 10.1053/j.gastro.2022.04.044. Epub 2022 Apr 29. Gastroenterology. 2022. PMID: 35500618 No abstract available.
References
-
- Rudnick DA, Perlmutter DH. Alpha-1-Antitrypsin Deficiency. In: Arias HJA Irwin M., Boyer James L., Cohen David E., Thorgeirsson Snorri, Shafritz David A., Wolkoff Allan W., ed. The Liver: Biology and Pathobiology. 6th ed: Wiley-Blackwell, 2020.
-
- Rudnick DA, Perlmutter DH. Alpha1-Antitrypsin Deficiency. In: Suchy RS F, Balistreri W, Bezerra J, Mack C, Shneider B, ed. Liver Disease in Children. 5th ed: Cambridge University Press, 2021:417–436.
-
- Rudnick DA, Perlmutter DH. Alpha-1-antitrypsin deficiency: a new paradigm for hepatocellular carcinoma in genetic liver disease. [Review] [49 refs]. Hepatology 2005;42:514–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
