

BTK inhibition limits B-cell-T-cell interaction through modulation of B-cell metabolism: implications for multiple sclerosis therapy
- PMID: 35303161
- PMCID: PMC8960592
- DOI: 10.1007/s00401-022-02411-w
BTK inhibition limits B-cell-T-cell interaction through modulation of B-cell metabolism: implications for multiple sclerosis therapy
Abstract
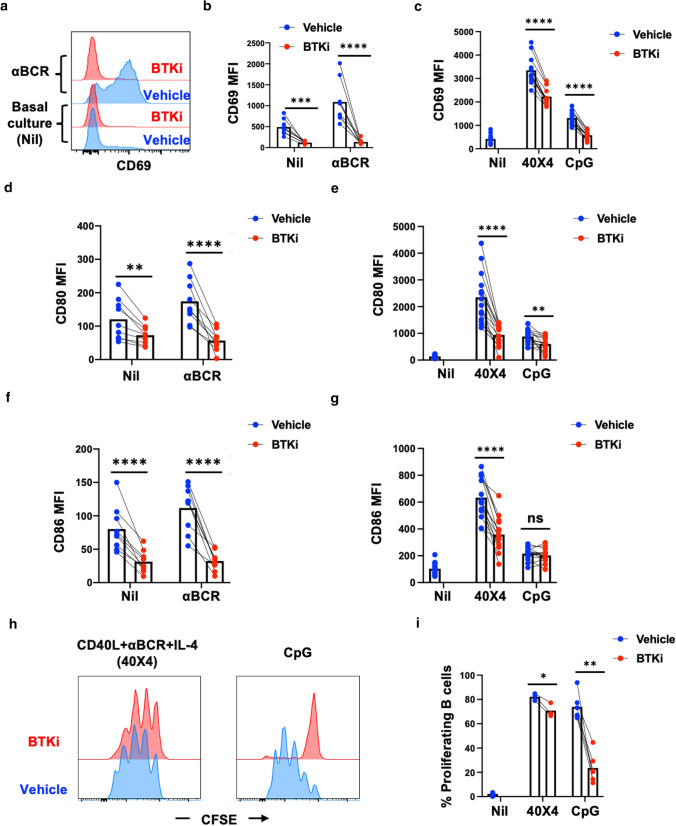
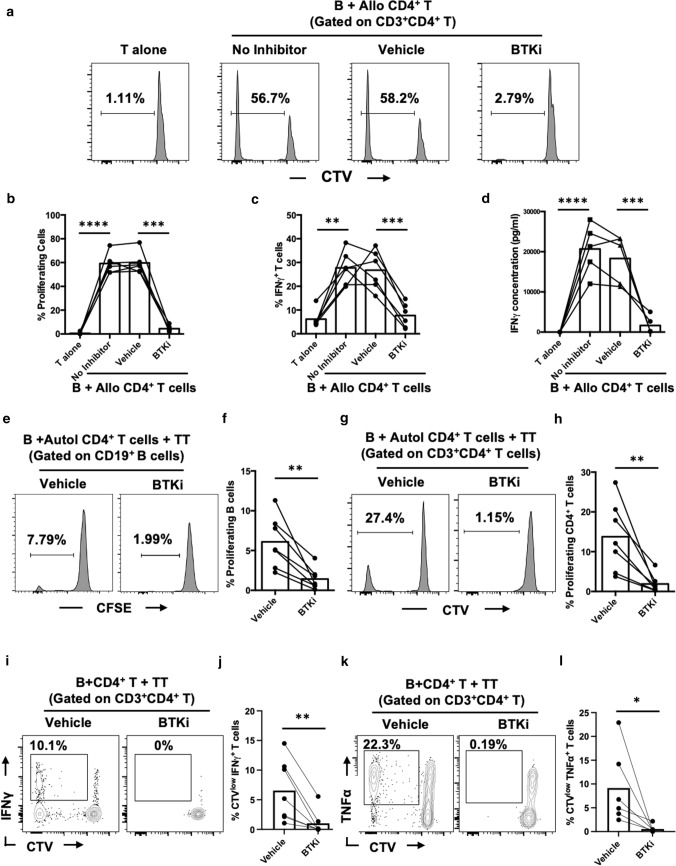
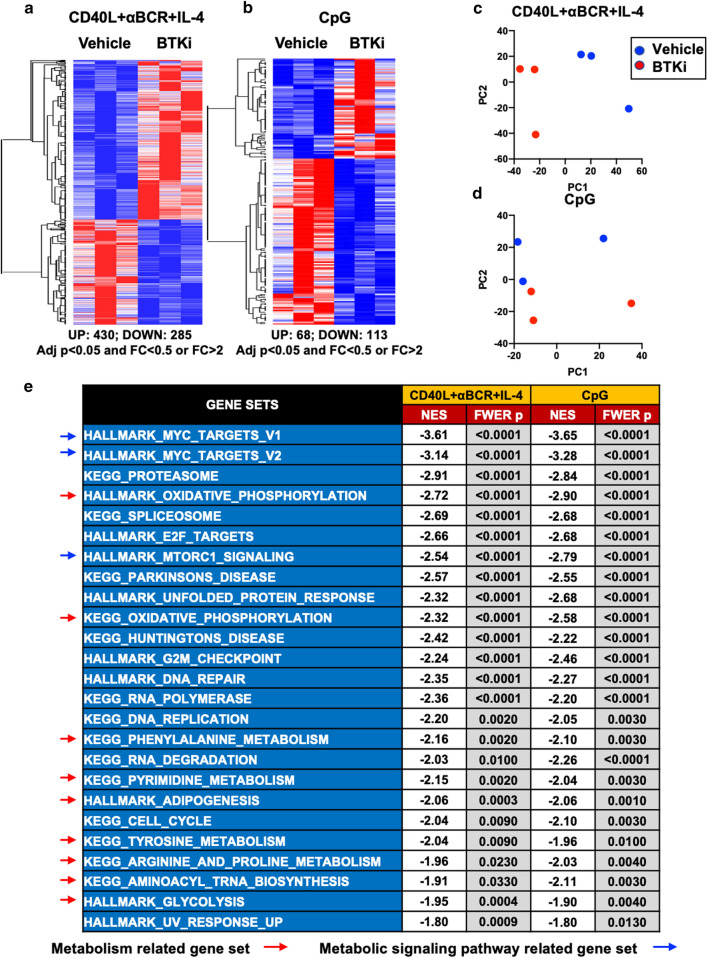
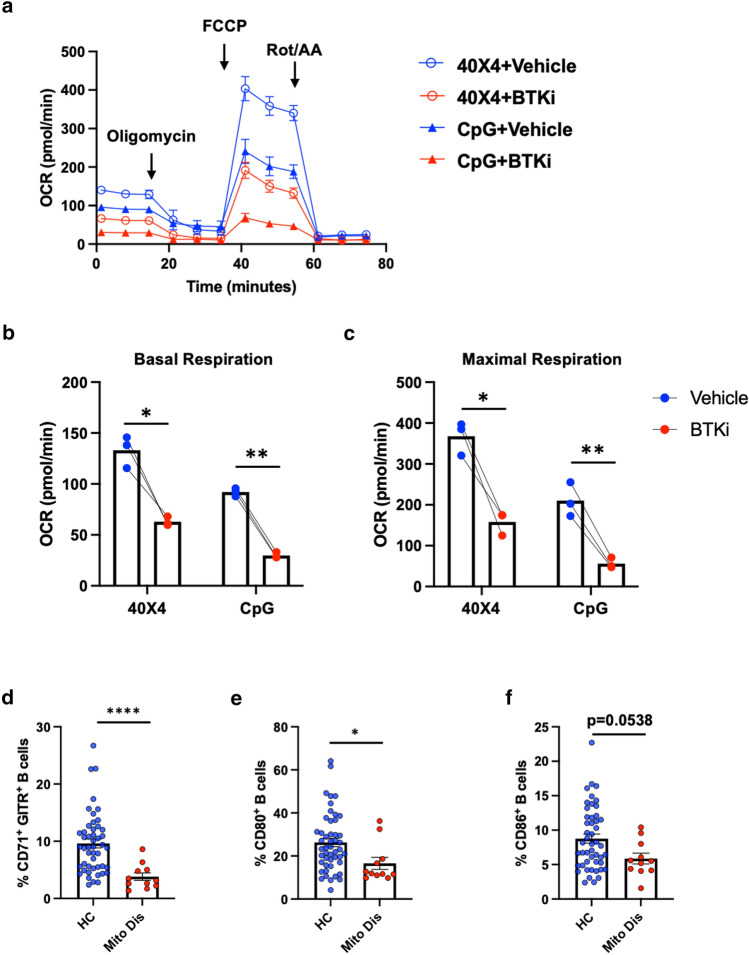
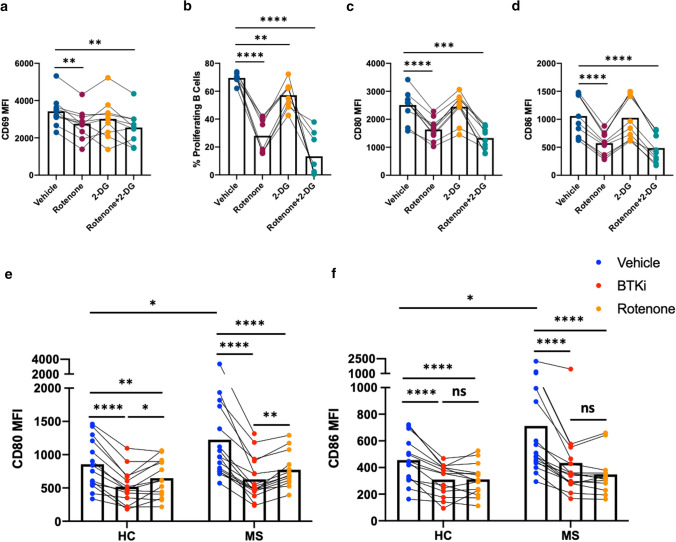
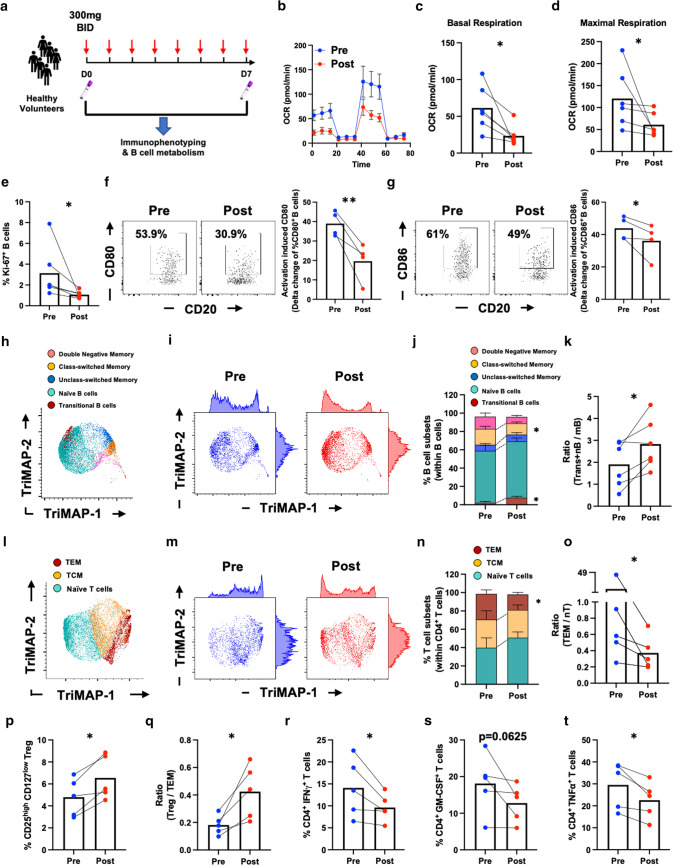
Inhibition of Bruton's Tyrosine Kinase (BTKi) is now viewed as a promising next-generation B-cell-targeting therapy for autoimmune diseases including multiple sclerosis (MS). Surprisingly little is known; however, about how BTKi influences MS disease-implicated functions of B cells. Here, we demonstrate that in addition to its expected impact on B-cell activation, BTKi attenuates B-cell:T-cell interactions via a novel mechanism involving modulation of B-cell metabolic pathways which, in turn, mediates an anti-inflammatory modulation of the B cells. In vitro, BTKi, as well as direct inhibition of B-cell mitochondrial respiration (but not glycolysis), limit the B-cell capacity to serve as APC to T cells. The role of metabolism in the regulation of human B-cell responses is confirmed when examining B cells of rare patients with mitochondrial respiratory chain mutations. We further demonstrate that both BTKi and metabolic modulation ex vivo can abrogate the aberrant activation and costimulatory molecule expression of B cells of untreated MS patients. Finally, as proof-of-principle in a Phase 1 study of healthy volunteers, we confirm that in vivo BTKi treatment reduces circulating B-cell mitochondrial respiration, diminishes their activation-induced expression of costimulatory molecules, and mediates an anti-inflammatory shift in the B-cell responses which is associated with an attenuation of T-cell pro-inflammatory responses. These data collectively elucidate a novel non-depleting mechanism by which BTKi mediates its effects on disease-implicated B-cell responses and reveals that modulating B-cell metabolism may be a viable therapeutic approach to target pro-inflammatory B cells.
Keywords: Autoimmune diseases; B cells; Bruton’s tyrosine kinase (BTK); Co-stimulatory molecules; Cytokines; Immunometabolism.
© 2022. The Author(s).
Conflict of interest statement
H.T, B.T.H and M.M are current employees of Biogen, MA. J.B was a former employee of Biogen and current employee of Abata Therapeutics. N.R receives salary support from the Jeffrey Modell Foundation. Amit Bar-Or has participated as a speaker in meetings sponsored by and received consulting fees and/or grant support from: Accure, Atara Biotherapeutics, Biogen, BMS/Celgene/Receptos, GlaxoSmithKline, Gossamer, Janssen/Actelion, Medimmune, Merck/EMD Serono, Novartis, Roche/Genentech, Sanofi-Genzyme.
Figures
Comment in
-
MS drug alters B cell metabolism.Nat Rev Neurol. 2022 May;18(5):252. doi: 10.1038/s41582-022-00655-4. Nat Rev Neurol. 2022. PMID: 35379952 No abstract available.
References
-
- Bame E, Tang H, Burns JC, Arefayene M, Michelsen K, Ma B, Marx I, Prince R, Roach AM, Poreci U, et al. Next-generation Bruton's tyrosine kinase inhibitor BIIB091 selectively and potently inhibits B cell and Fc receptor signaling and downstream functions in B cells and myeloid cells. Clin Transl Immunol. 2021;10:e1295. doi: 10.1002/cti2.1295. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
