

Astrocytic GABA transporter controls sleep by modulating GABAergic signaling in Drosophila circadian neurons
- PMID: 35303417
- PMCID: PMC9090989
- DOI: 10.1016/j.cub.2022.02.066
Astrocytic GABA transporter controls sleep by modulating GABAergic signaling in Drosophila circadian neurons
Abstract
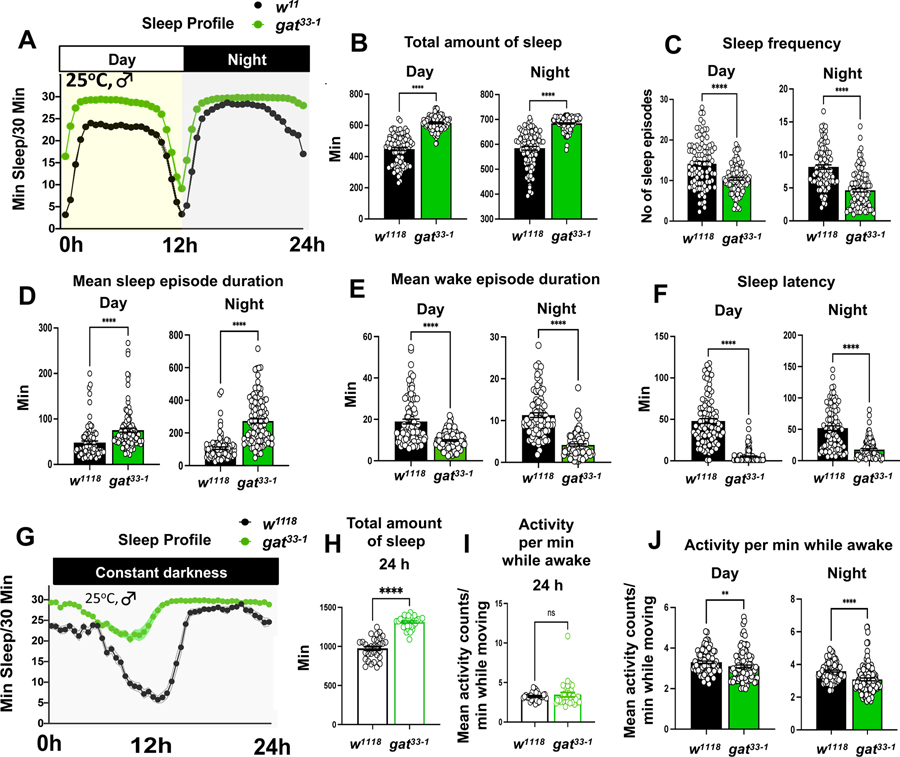
A precise balance between sleep and wakefulness is essential to sustain a good quality of life and optimal brain function. GABA is known to play a key and conserved role in sleep control, and GABAergic tone should, therefore, be tightly controlled in sleep circuits. Here, we examined the role of the astrocytic GABA transporter (GAT) in sleep regulation using Drosophila melanogaster. We found that a hypomorphic gat mutation (gat33-1) increased sleep amount, decreased sleep latency, and increased sleep consolidation at night. Interestingly, sleep defects were suppressed when gat33-1 was combined with a mutation disrupting wide-awake (wake), a gene that regulates the cell-surface levels of the GABAA receptor resistance to dieldrin (RDL) in the wake-promoting large ventral lateral neurons (l-LNvs). Moreover, RNAi knockdown of rdl and its modulators dnlg4 and wake in these circadian neurons also suppressed gat33-1 sleep phenotypes. Brain immunohistochemistry showed that GAT-expressing astrocytes were located near RDL-positive l-LNv cell bodies and dendritic processes. We concluded that astrocytic GAT decreases GABAergic tone and RDL activation in arousal-promoting LNvs, thus determining proper sleep amount and quality in Drosophila.
Keywords: GABA; GAT; astrocytes; large ventral lateral neurons; sleep.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Similar articles
-
Drosophila neuroligin 4 regulates sleep through modulating GABA transmission.J Neurosci. 2013 Sep 25;33(39):15545-54. doi: 10.1523/JNEUROSCI.0819-13.2013. J Neurosci. 2013. PMID: 24068821 Free PMC article.
-
PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit.Neuron. 2008 Nov 26;60(4):672-82. doi: 10.1016/j.neuron.2008.10.042. Neuron. 2008. PMID: 19038223 Free PMC article.
-
The GABA(A) receptor RDL acts in peptidergic PDF neurons to promote sleep in Drosophila.Curr Biol. 2009 Mar 10;19(5):386-90. doi: 10.1016/j.cub.2009.01.040. Epub 2009 Feb 19. Curr Biol. 2009. PMID: 19230663 Free PMC article.
-
GABAergic modulation of sleep-wake states.Pharmacol Ther. 2023 Sep;249:108505. doi: 10.1016/j.pharmthera.2023.108505. Epub 2023 Aug 3. Pharmacol Ther. 2023. PMID: 37541595 Review.
-
Circadian Rhythms and Sleep in Drosophila melanogaster.Genetics. 2017 Apr;205(4):1373-1397. doi: 10.1534/genetics.115.185157. Genetics. 2017. PMID: 28360128 Free PMC article. Review.
Cited by
-
Astrocytes at the heart of sleep: from genes to network dynamics.Cell Mol Life Sci. 2025 May 21;82(1):207. doi: 10.1007/s00018-025-05671-3. Cell Mol Life Sci. 2025. PMID: 40397158 Free PMC article. Review.
-
A transporter's doom or destiny: SLC6A1 in health and disease, novel molecular targets and emerging therapeutic prospects.Front Mol Neurosci. 2024 Aug 29;17:1466694. doi: 10.3389/fnmol.2024.1466694. eCollection 2024. Front Mol Neurosci. 2024. PMID: 39268250 Free PMC article. Review.
-
Role of astrocytes in sleep deprivation: accomplices, resisters, or bystanders?Front Cell Neurosci. 2023 Jun 26;17:1188306. doi: 10.3389/fncel.2023.1188306. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37435045 Free PMC article. Review.
-
Contributions of Gamma-Aminobutyric Acid (GABA) Produced by Lactic Acid Bacteria on Food Quality and Human Health: Current Applications and Future Prospects.Foods. 2024 Aug 1;13(15):2437. doi: 10.3390/foods13152437. Foods. 2024. PMID: 39123629 Free PMC article. Review.
-
Glial Regulation of Circuit Wiring, Firing, and Expiring in the Drosophila Central Nervous System.Cold Spring Harb Perspect Biol. 2024 Dec 2;16(12):a041347. doi: 10.1101/cshperspect.a041347. Cold Spring Harb Perspect Biol. 2024. PMID: 38565270 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
