

A novel role for GalNAc-T2 dependent glycosylation in energy homeostasis
- PMID: 35304331
- PMCID: PMC9019398
- DOI: 10.1016/j.molmet.2022.101472
A novel role for GalNAc-T2 dependent glycosylation in energy homeostasis
Abstract
Objective: GALNT2, encoding polypeptide N-acetylgalactosaminyltransferase 2 (GalNAc-T2), was initially discovered as a regulator of high-density lipoprotein metabolism. GalNAc-T2 is known to exert these effects through post-translational modification, i.e., O-linked glycosylation of secreted proteins with established roles in plasma lipid metabolism. It has recently become clear that loss of GALNT2 in rodents, cattle, nonhuman primates, and humans should be regarded as a novel congenital disorder of glycosylation that affects development and body weight. The role of GALNT2 in metabolic abnormalities other than plasma lipids, including insulin sensitivity and energy homeostasis, is poorly understood.
Methods: GWAS data from the UK Biobank was used to study variation in the GALNT2 locus beyond changes in high-density lipoprotein metabolism. Experimental data were obtained through studies in Galnt2-/- mice and wild-type littermates on both control and high-fat diet.
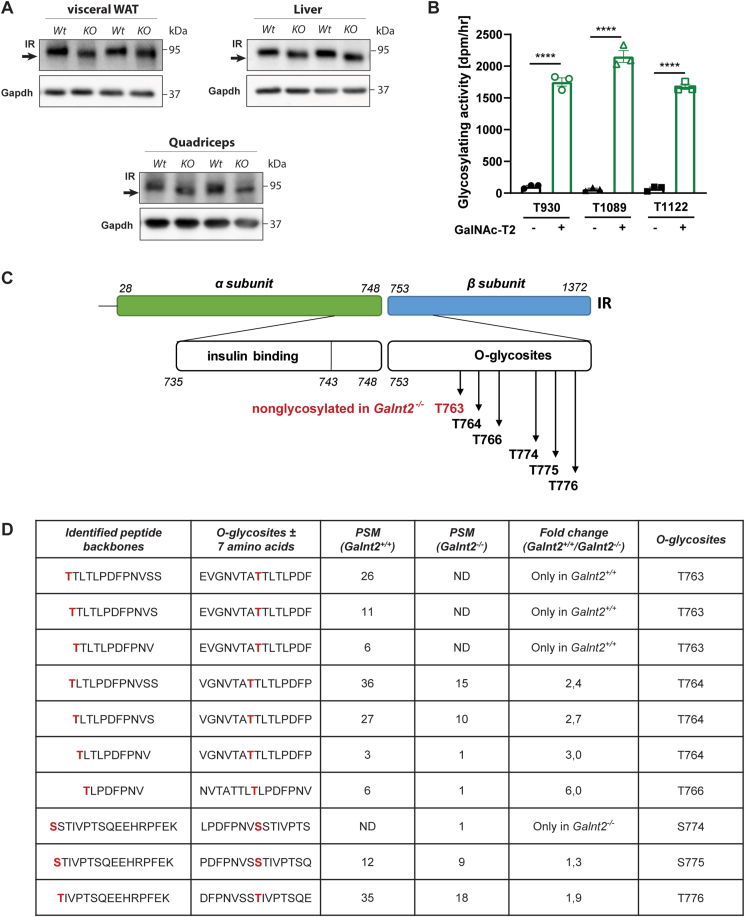
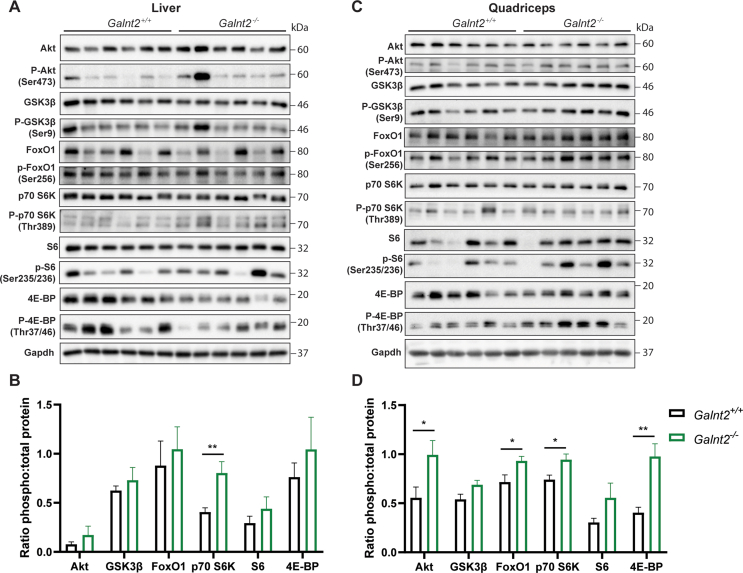
Results: First, we uncovered associations between GALNT2 gene variation, adiposity, and body mass index in humans. In mice, we identify the insulin receptor as a novel substrate of GalNAc-T2 and demonstrate that Galnt2-/- mice exhibit decreased adiposity, alterations in insulin signaling and a shift in energy substrate utilization in the inactive phase.
Conclusions: This study identifies a novel role for GALNT2 in energy homeostasis, and our findings suggest that the local effects of GalNAc-T2 are mediated through posttranslational modification of the insulin receptor.
Keywords: Adipose tissue; Energy metabolism; Genetic disorder; Glycosylation; Insulin signaling.
Copyright © 2022 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures














References
-
- Schjoldager K.T.B.G., Vester-Christensen M.B., Bennett E.P., Levery S.B., Schwientek T., Yin W., et al. O-glycosylation modulates proprotein convertase activation of angiopoietin-like protein 3: possible role of polypeptide GalNAc-transferase-2 in regulation of concentrations of plasma lipids. Journal of Biological Chemistry. 2010;285(47):36293–36303. doi: 10.1074/jbc.M110.156950. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
