

Analysis tools for single-monomer measurements of self-assembly processes
- PMID: 35304498
- PMCID: PMC8933434
- DOI: 10.1038/s41598-022-08245-6
Analysis tools for single-monomer measurements of self-assembly processes
Abstract
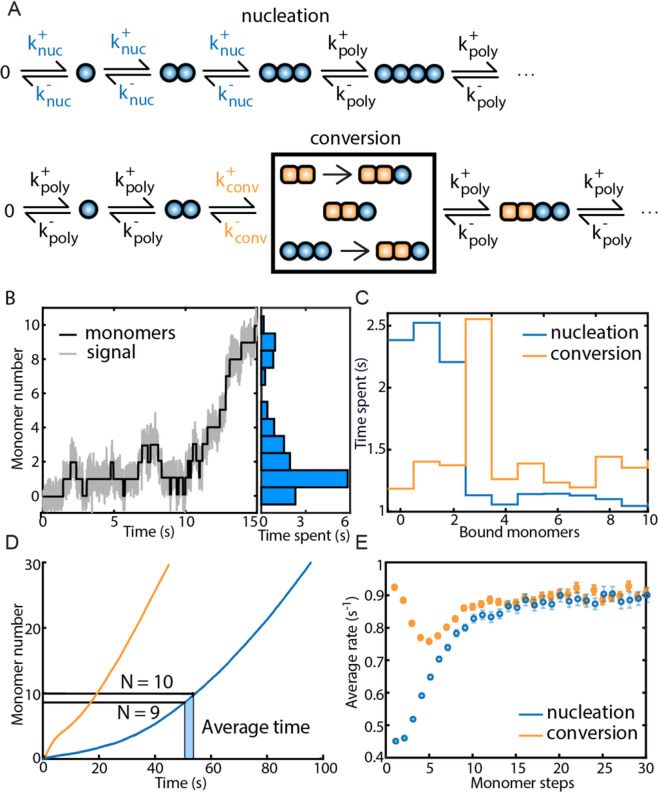
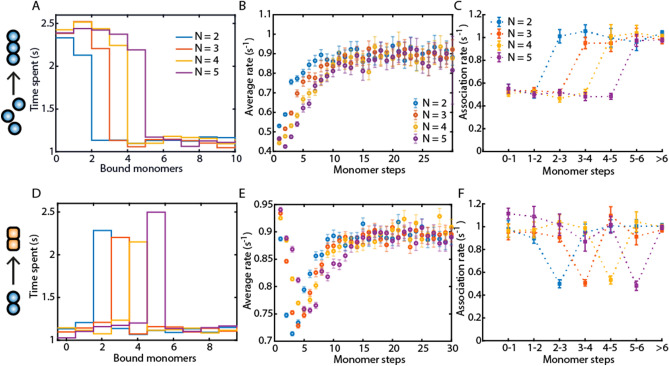
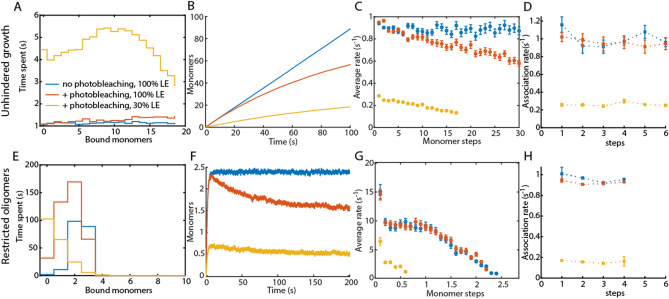
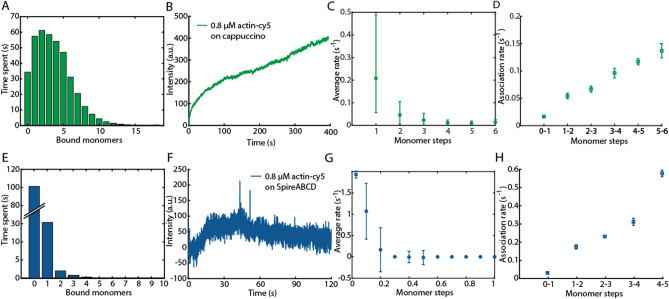
Protein assembly plays an important role throughout all phyla of life, both physiologically and pathologically. In particular, aggregation and polymerization of proteins are key-strategies that regulate cellular function. In recent years, methods to experimentally study the assembly process on a single-molecule level have been developed. This progress concomitantly has triggered the question of how to analyze this type of single-filament data adequately and what experimental conditions are necessary to allow a meaningful interpretation of the analysis. Here, we developed two analysis methods for single-filament data: the visitation analysis and the average-rate analysis. We benchmarked and compared both approaches with the classic dwell-time-analysis frequently used to study microscopic association and dissociation rates. In particular, we tested the limitations of each analysis method along the lines of the signal-to-noise ratio, the sampling rate, and the labeling efficiency and bleaching rate of the fluorescent dyes used in single-molecule fluorescence experiments. Finally, we applied our newly developed methods to study the monomer assembly of actin at the single-molecule-level in the presence of the class II nucleator Cappuccino and the WH2 repeats of Spire. For Cappuccino, our data indicated fast elongation circumventing a nucleation phase whereas, for Spire, we found that the four WH2 motifs are not sufficient to promote de novo nucleation of actin.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Drosophila Spire is an actin nucleation factor.Nature. 2005 Jan 27;433(7024):382-8. doi: 10.1038/nature03241. Nature. 2005. PMID: 15674283
-
The WH2 Domain and Actin Nucleation: Necessary but Insufficient.Trends Biochem Sci. 2016 Jun;41(6):478-490. doi: 10.1016/j.tibs.2016.03.004. Epub 2016 Apr 5. Trends Biochem Sci. 2016. PMID: 27068179 Free PMC article. Review.
-
Actin nucleation: spire - actin nucleator in a class of its own.Curr Biol. 2005 Apr 26;15(8):R305-8. doi: 10.1016/j.cub.2005.04.004. Curr Biol. 2005. PMID: 15854898 Review.
-
Spire stimulates nucleation by Cappuccino and binds both ends of actin filaments.Mol Biol Cell. 2020 Feb 15;31(4):273-286. doi: 10.1091/mbc.E19-09-0550. Epub 2019 Dec 26. Mol Biol Cell. 2020. PMID: 31877067 Free PMC article.
-
Analysis of the function of Spire in actin assembly and its synergy with formin and profilin.Mol Cell. 2007 Nov 30;28(4):555-68. doi: 10.1016/j.molcel.2007.09.018. Mol Cell. 2007. PMID: 18042452
Cited by
-
Zero-mode waveguides visualize the first steps during gelsolin-mediated actin filament formation.Biophys J. 2022 Jan 18;121(2):327-335. doi: 10.1016/j.bpj.2021.12.011. Epub 2021 Dec 9. Biophys J. 2022. PMID: 34896371 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
