

Molecular mechanism of ethanol fermentation inhibition via protein tyrosine nitration of pyruvate decarboxylase by reactive nitrogen species in yeast
- PMID: 35304512
- PMCID: PMC8933545
- DOI: 10.1038/s41598-022-08568-4
Molecular mechanism of ethanol fermentation inhibition via protein tyrosine nitration of pyruvate decarboxylase by reactive nitrogen species in yeast
Abstract
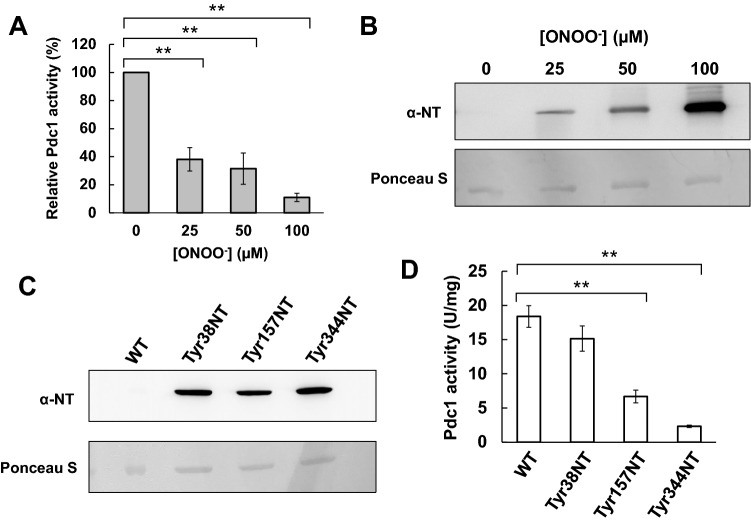
Protein tyrosine nitration (PTN), in which tyrosine (Tyr) residues on proteins are converted into 3-nitrotyrosine (NT), is one of the post-translational modifications mediated by reactive nitrogen species (RNS). Many recent studies have reported that PTN contributed to signaling systems by altering the structures and/or functions of proteins. This study aimed to investigate connections between PTN and the inhibitory effect of nitrite-derived RNS on fermentation ability using the yeast Saccharomyces cerevisiae. The results indicated that RNS inhibited the ethanol production of yeast cells with increased intracellular pyruvate content. We also found that RNS decreased the activities of pyruvate decarboxylase (PDC) as a critical enzyme involved in ethanol production. Our proteomic analysis revealed that the main PDC isozyme Pdc1 underwent the PTN modification at Tyr38, Tyr157, and Tyr344. The biochemical analysis using the recombinant purified Pdc1 enzyme indicated that PTN at Tyr157 or Tyr344 significantly reduced the Pdc1 activity. Interestingly, the substitution of Tyr157 or Tyr344 to phenylalanine, which is no longer converted into NT, recovered the ethanol production under the RNS treatment conditions. These findings suggest that nitrite impairs the fermentation ability of yeast by inhibiting the Pdc1 activity via its PTN modification at Tyr157 and Tyr344 of Pdc1.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Brüne B, Meβmer UK, Sandau K. The role of nitric oxide in cell injury. Toxicol. Lett. 1995;82–83:233–237. - PubMed
-
- Schuman EM, Madison DV. A requirement for the intercellular messenger nitric oxide in long-term potentiation. Science. 1991;254:1503–1506. - PubMed
-
- Pozdnyakov N, Lloyd A, Reddy VN, Sitaramayya A. Nitric oxide-regulated endogenous ADP-ribosylation of rod outer segment proteins. Biochem. Biophys. Res. Commun. 1993;192:610–615. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
