

Ejection of damaged mitochondria and their removal by macrophages ensure efficient thermogenesis in brown adipose tissue
- PMID: 35305295
- PMCID: PMC9039922
- DOI: 10.1016/j.cmet.2022.02.016
Ejection of damaged mitochondria and their removal by macrophages ensure efficient thermogenesis in brown adipose tissue
Abstract
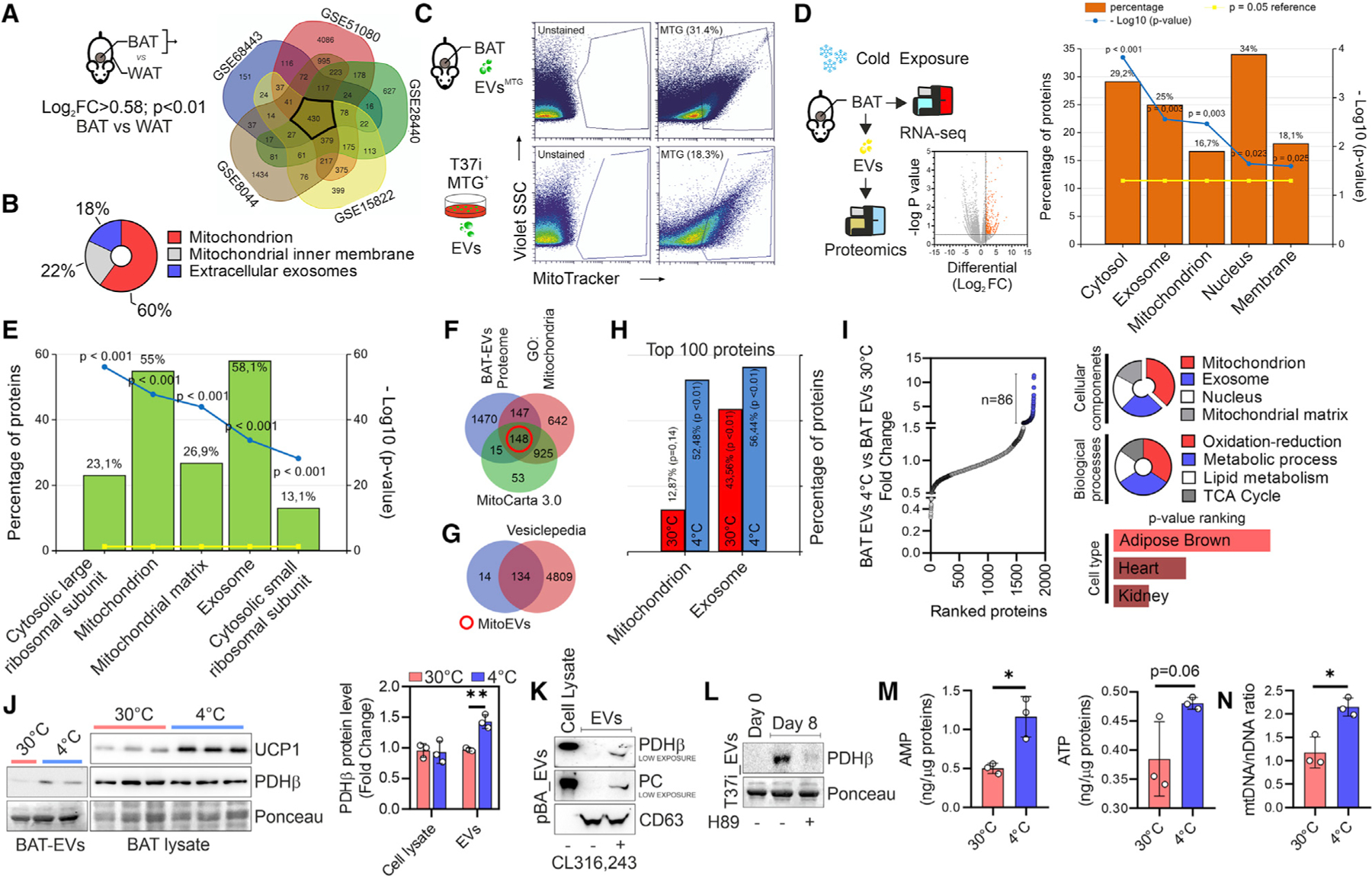
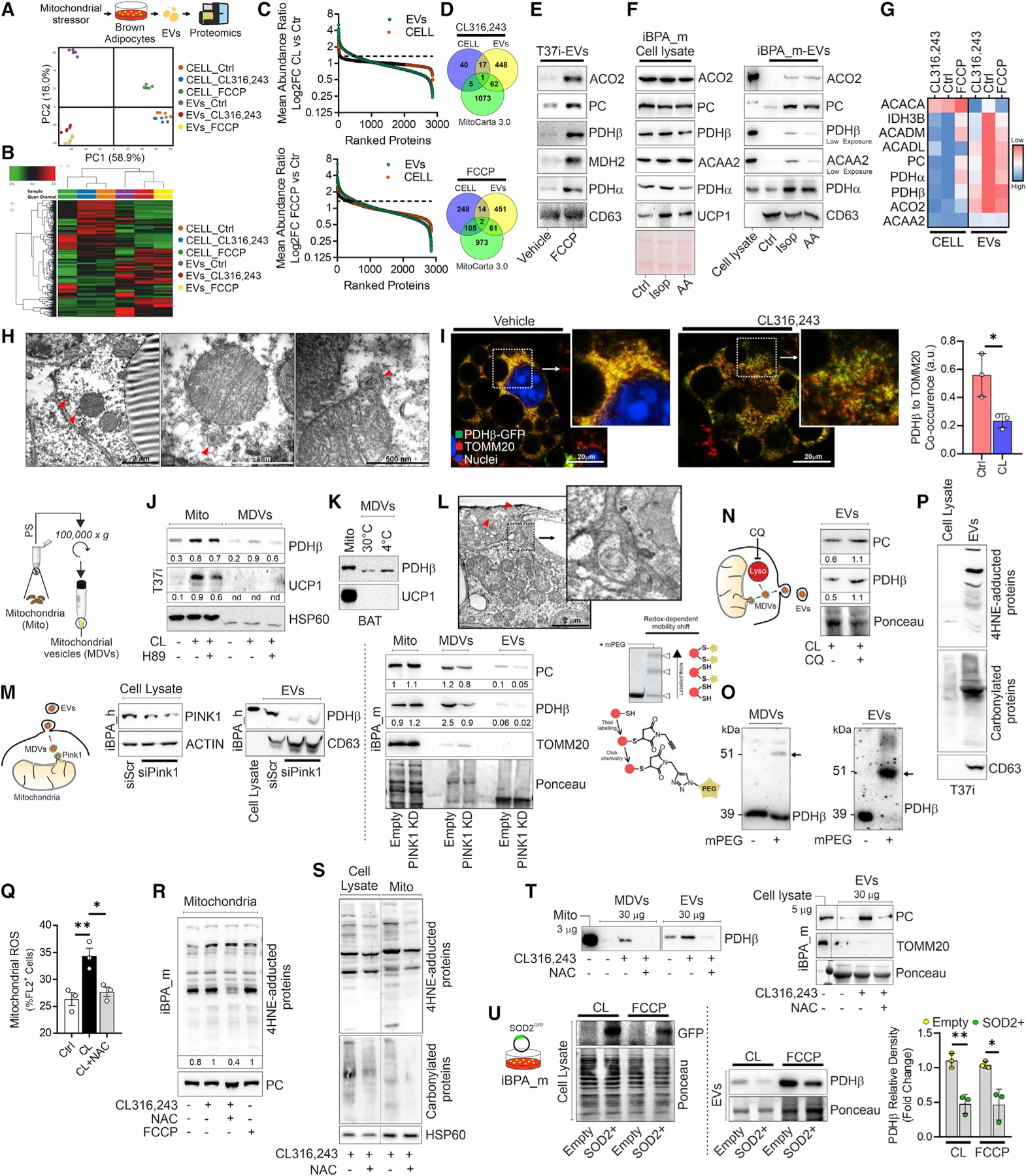
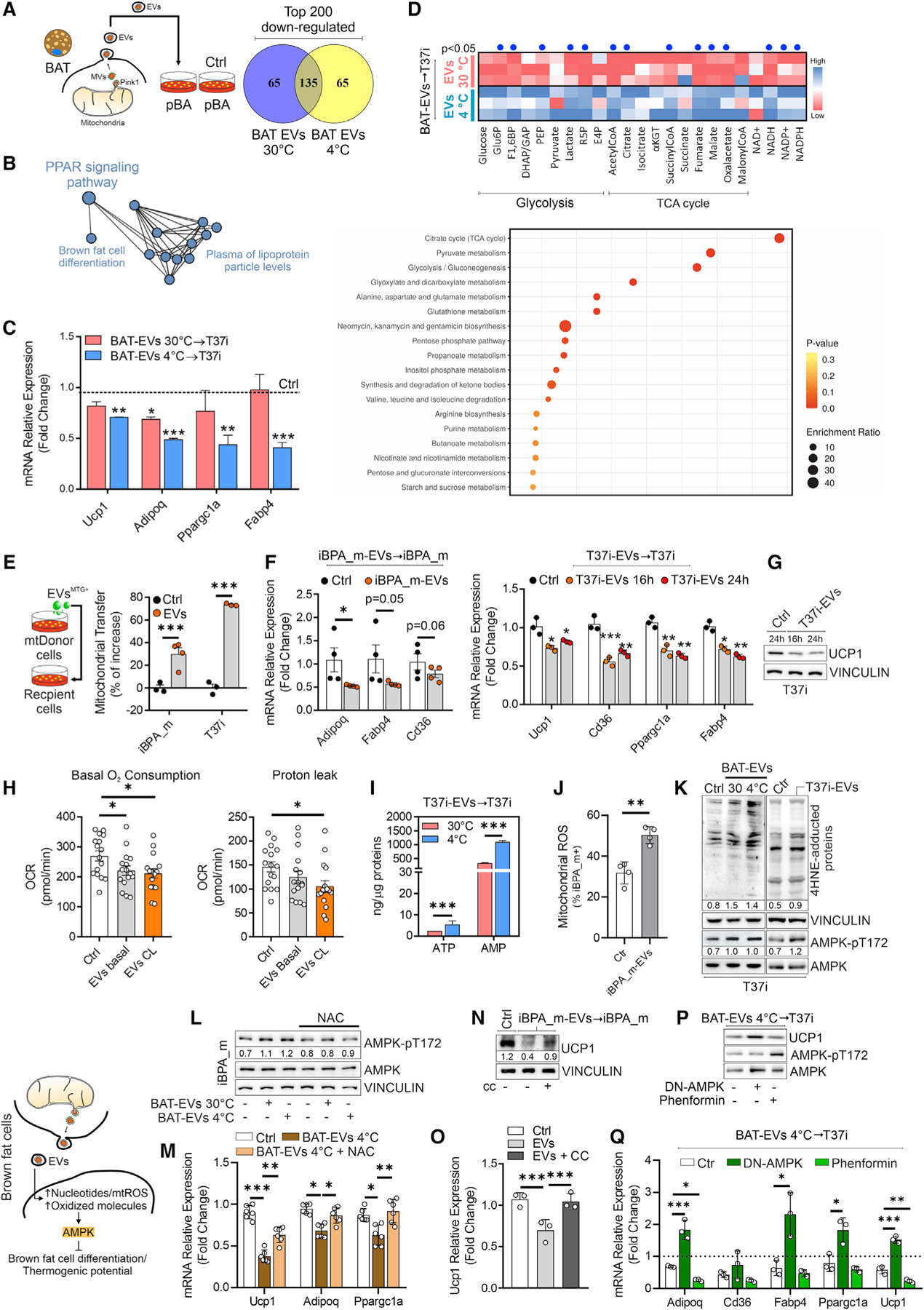
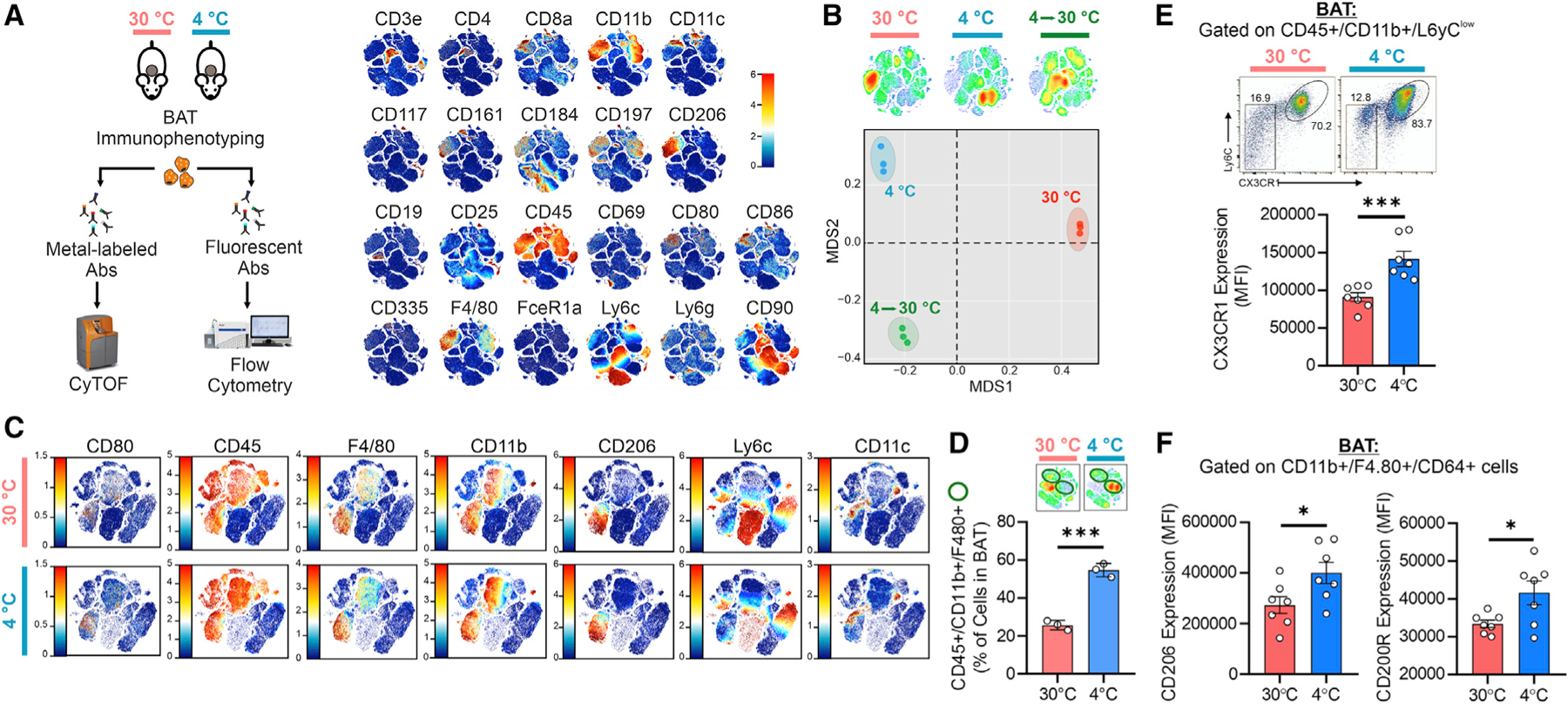
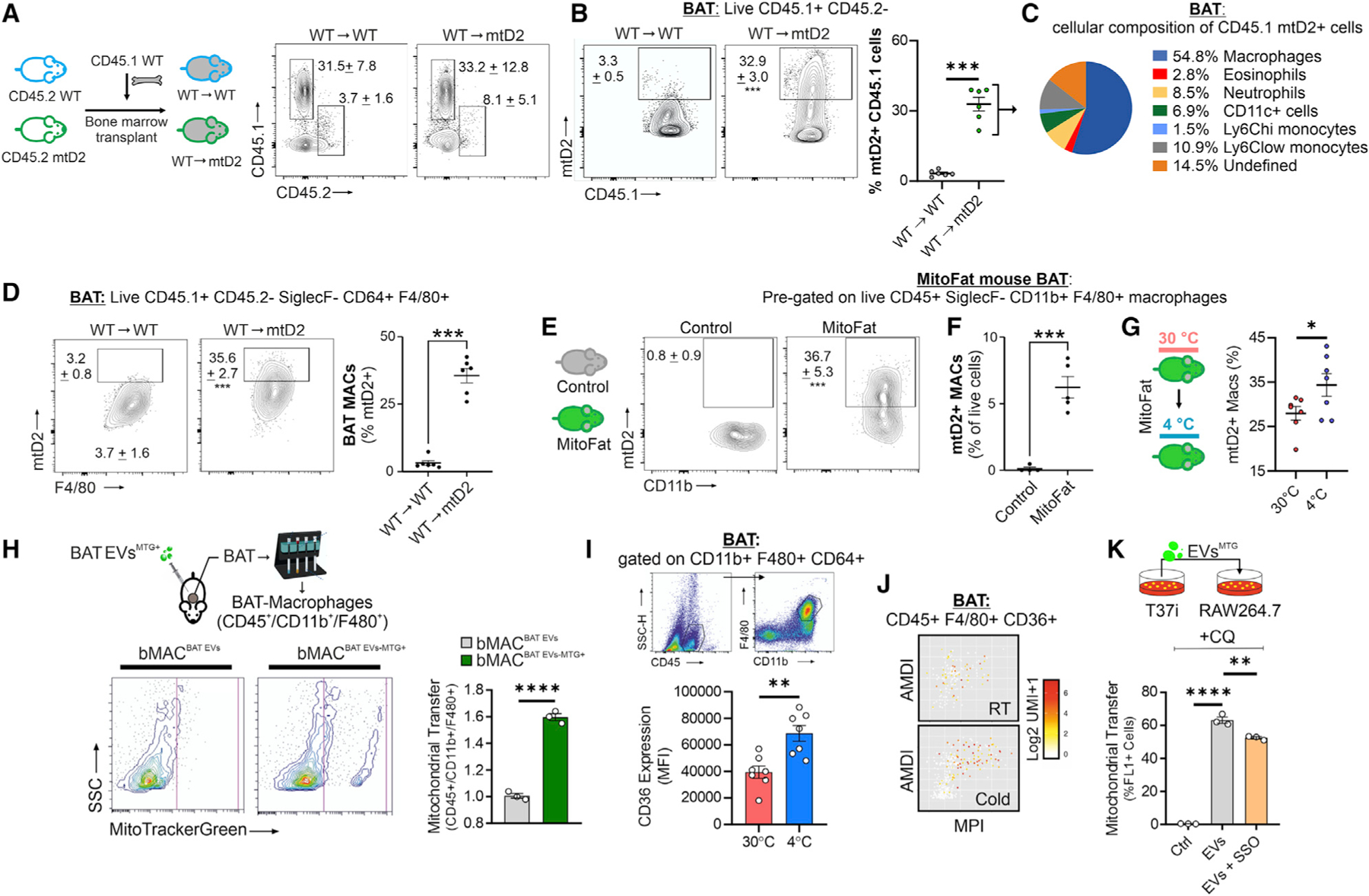
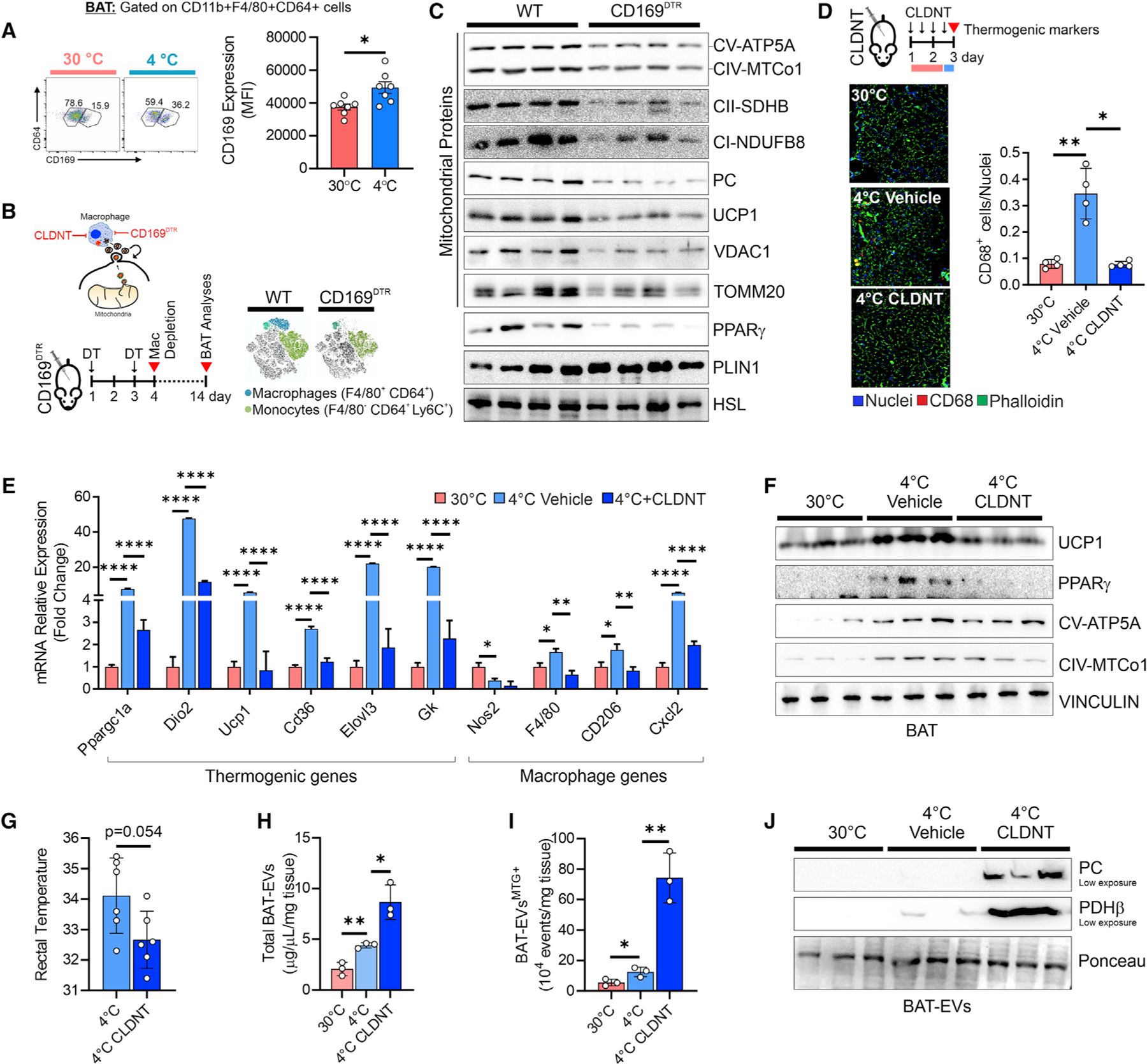
Recent findings have demonstrated that mitochondria can be transferred between cells to control metabolic homeostasis. Although the mitochondria of brown adipocytes comprise a large component of the cell volume and undergo reorganization to sustain thermogenesis, it remains unclear whether an intercellular mitochondrial transfer occurs in brown adipose tissue (BAT) and regulates adaptive thermogenesis. Herein, we demonstrated that thermogenically stressed brown adipocytes release extracellular vesicles (EVs) that contain oxidatively damaged mitochondrial parts to avoid failure of the thermogenic program. When re-uptaken by parental brown adipocytes, mitochondria-derived EVs reduced peroxisome proliferator-activated receptor-γ signaling and the levels of mitochondrial proteins, including UCP1. Their removal via the phagocytic activity of BAT-resident macrophages is instrumental in preserving BAT physiology. Depletion of macrophages in vivo causes the abnormal accumulation of extracellular mitochondrial vesicles in BAT, impairing the thermogenic response to cold exposure. These findings reveal a homeostatic role of tissue-resident macrophages in the mitochondrial quality control of BAT.
Keywords: adipose tissue; brown adipocytes; extracellular vesicles; homeostasis; immunometabolism; macrophages; mitochondria; mitochondrial quality control; thermogenesis.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.R.B. has a pending patent application related to mitochondria transfer. The other authors declare no competing interests.
Figures
Comment in
-
Mitochondrial transfer in BAT.Nat Rev Endocrinol. 2022 Jun;18(6):334. doi: 10.1038/s41574-022-00674-7. Nat Rev Endocrinol. 2022. PMID: 35393577 No abstract available.
References
-
- Aquilano K, Sciarretta F, Turchi R, Li BH, Rosina M, Ceci V, Guidobaldi G, Arena S, D’Ambrosio C, Audano M, et al. (2020). Low-protein/high-carbohydrate diet induces AMPK-dependent canonical and non-canonical thermogenesis in subcutaneous adipose tissue. Redox Biol 36, 101633. 10.1016/j.redox.2020.101633. - DOI - PMC - PubMed
