

Soluble ECM promotes organotypic formation in lung alveolar model
- PMID: 35306229
- PMCID: PMC9359416
- DOI: 10.1016/j.biomaterials.2022.121464
Soluble ECM promotes organotypic formation in lung alveolar model
Abstract
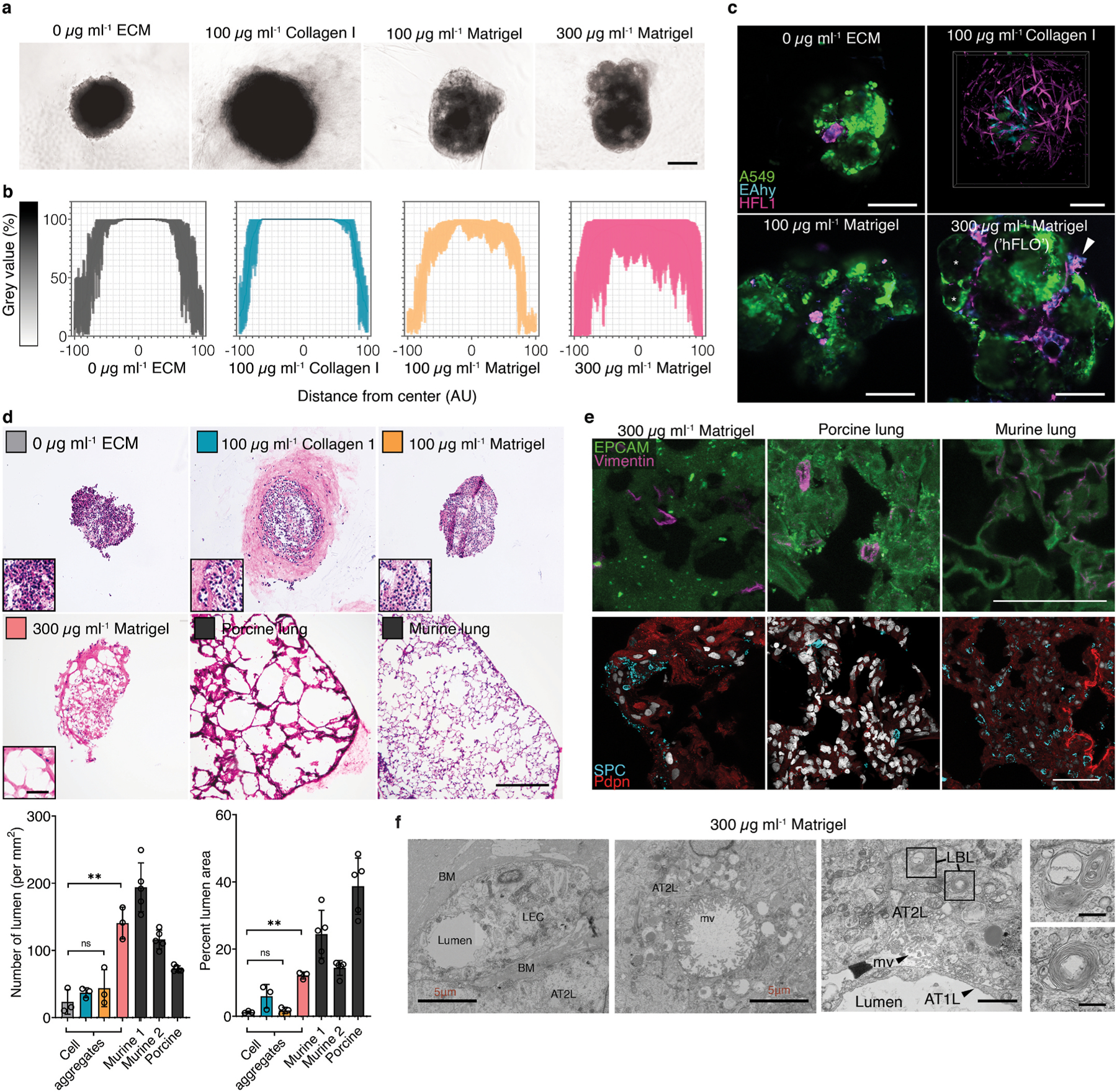
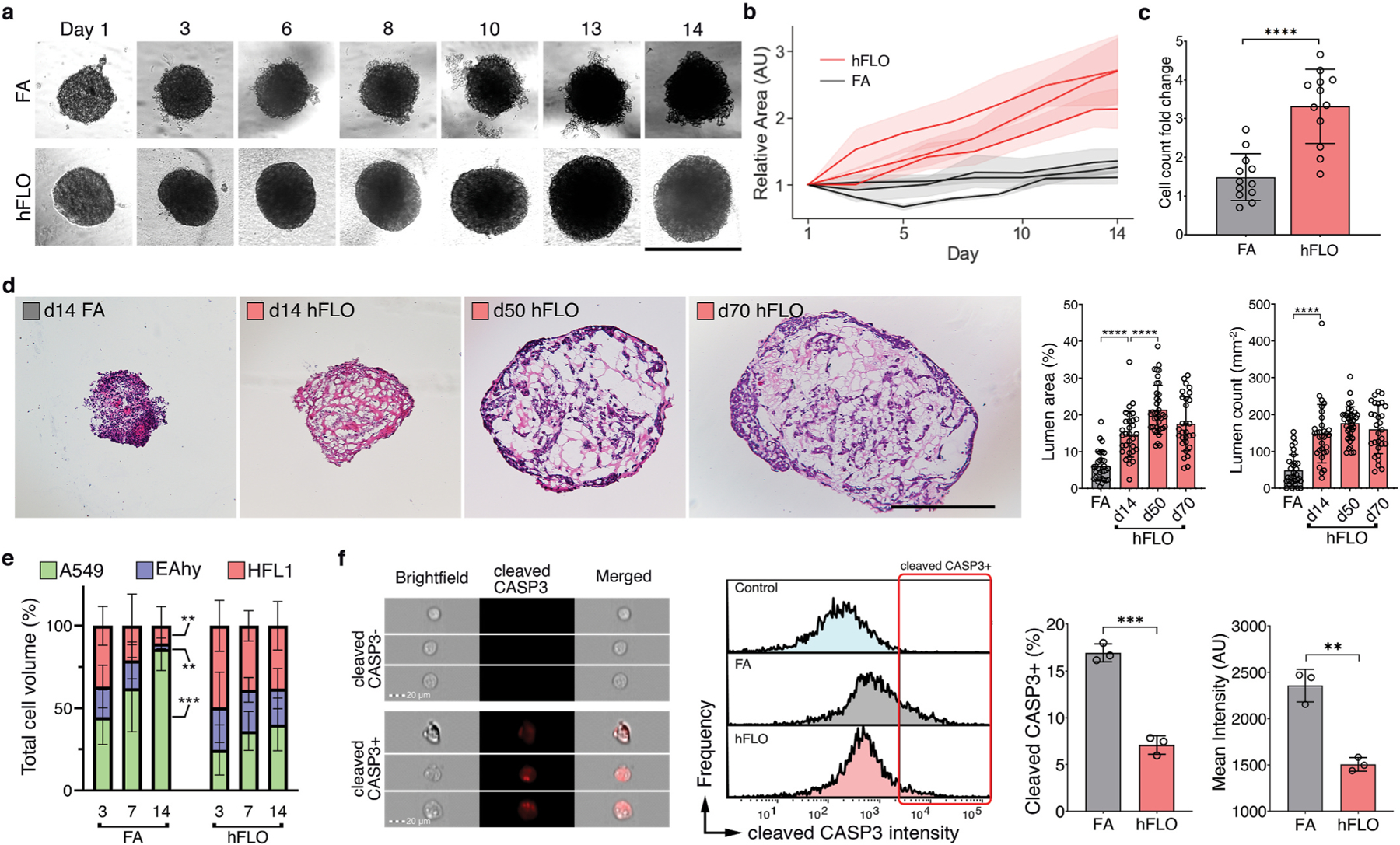
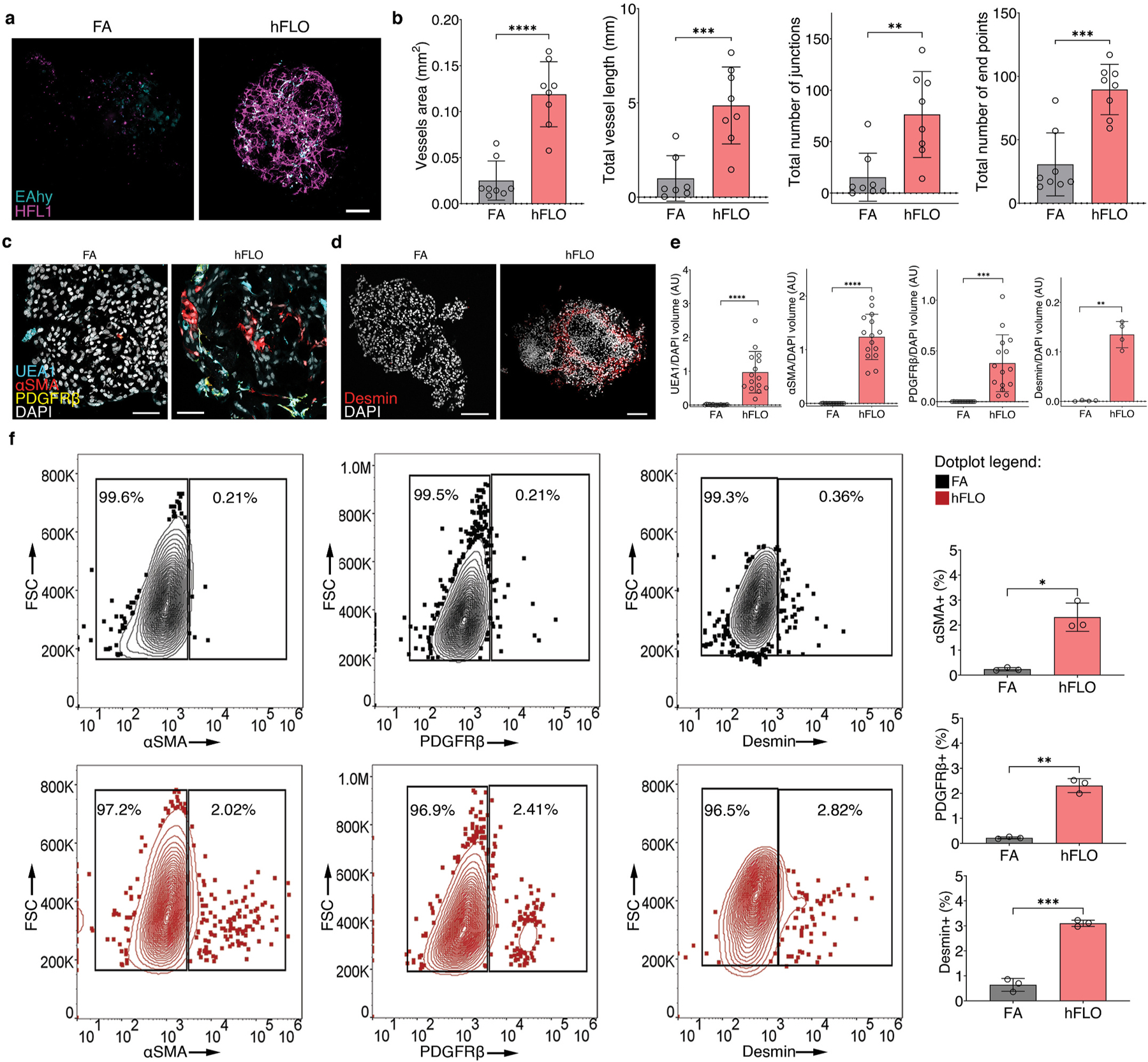
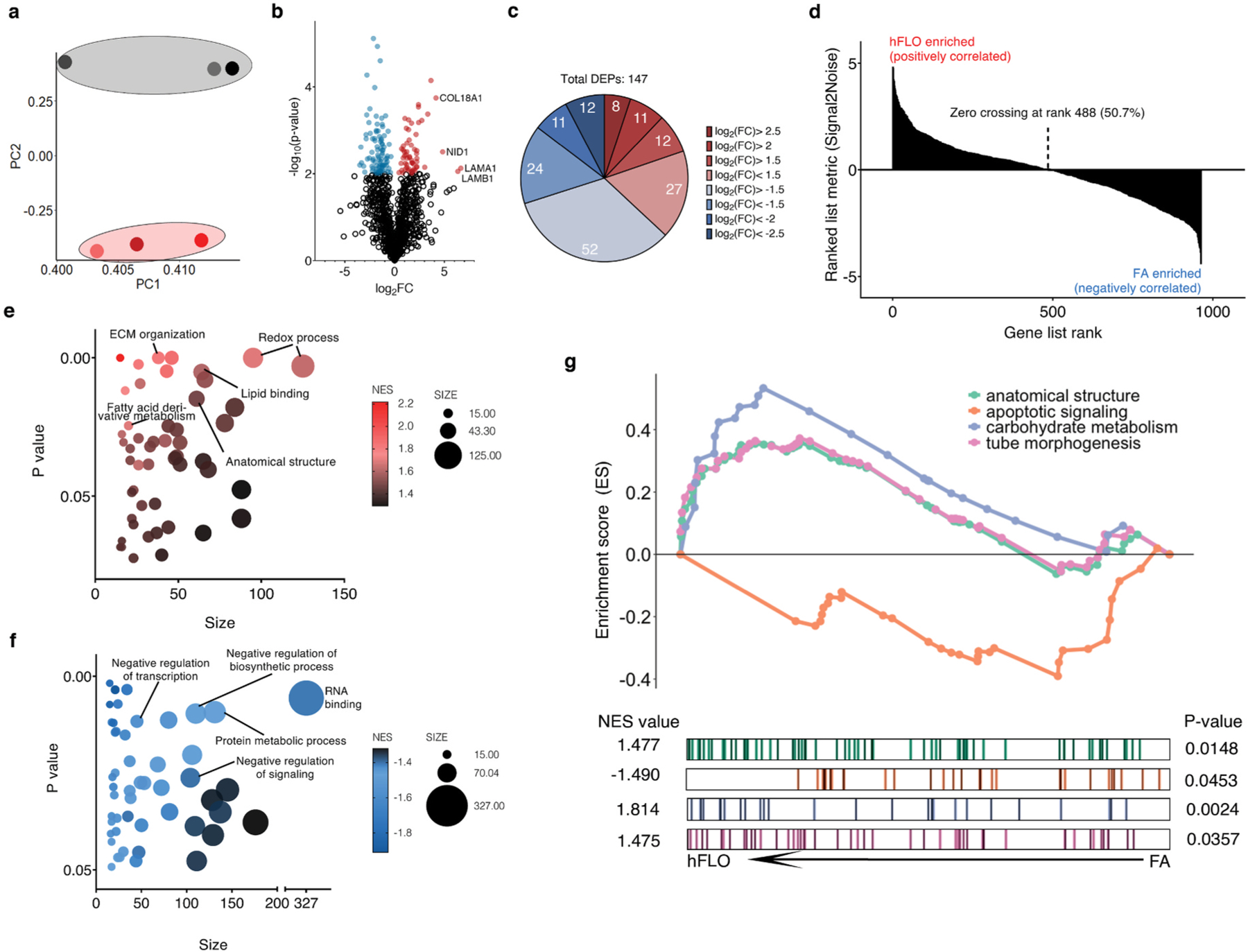
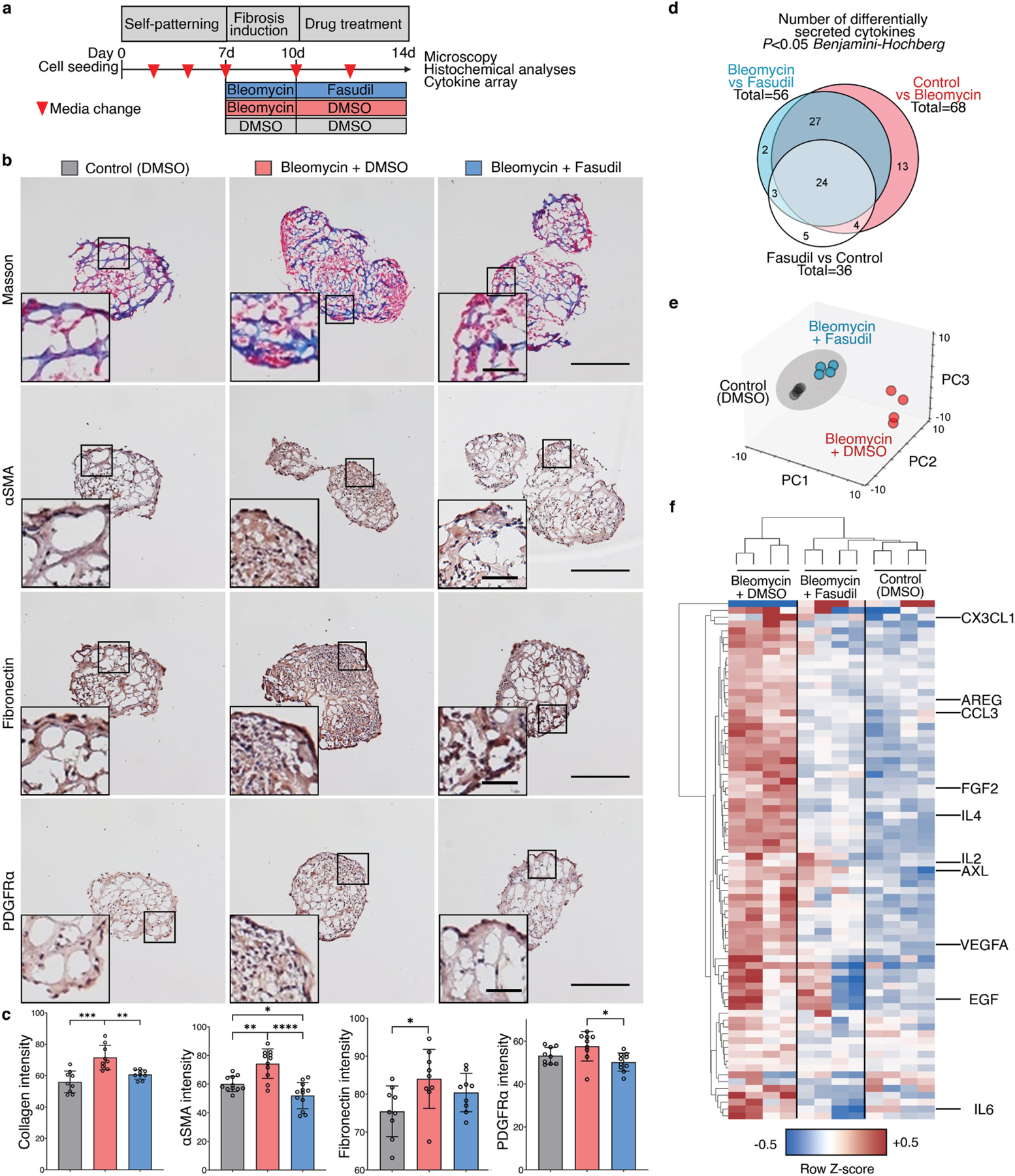
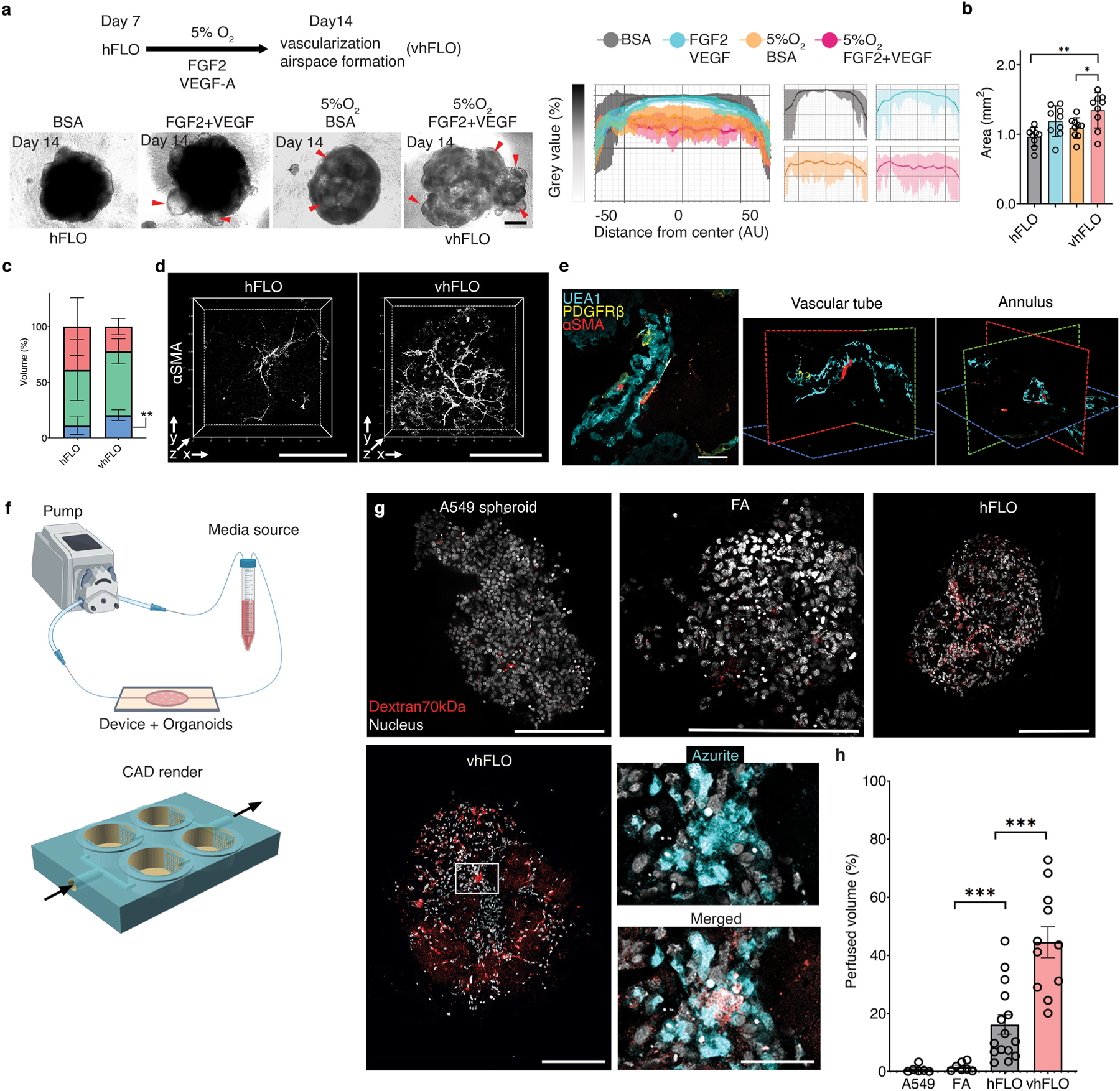
Micropatterned suspension culture creates consistently sized and shaped cell aggregates but has not produced organotypic structures from stable cells, thus restricting its use in accurate disease modeling. Here, we show that organotypic structure is achieved in hybrid suspension culture via supplementation of soluble extracellular matrix (ECM). We created a viable lung organoid from epithelial, endothelial, and fibroblast human stable cell lines in suspension culture. We demonstrate the importance of soluble ECM in organotypic patterning with the emergence of lumen-like structures with airspace showing feasible gas exchange units, formation of branching, perfusable vasculature, and long-term 70-day maintenance of lumen structure. Our results show a dependent relationship between enhanced fibronectin fibril assembly and the incorporation of ECM in the organoid. We successfully applied this technology in modeling lung fibrosis via bleomycin induction and test a potential antifibrotic drug in vitro while maintaining fundamental cell-cell interactions in lung tissue. Our human fluorescent lung organoid (hFLO) model represents features of pulmonary fibrosis which were ameliorated by fasudil treatment. We also demonstrate a 3D culture method with potential of creating organoids from mature cells, thus opening avenues for disease modeling and regenerative medicine, enhancing understanding of lung cell biology in health and lung disease.
Keywords: Extracellular matrix; Lung organoid; Pulmonary fibrosis; Suspension culture; Tissue engineering; Vascularization.
Copyright © 2022 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Jonard C. Valdoz and Pam M. Van Ry filed a provisional patent for “A Suspension-based 3D culture method for stable or primary cells and a fluorescent lung triculture organoid” (Provisional Pat. No. 63185423, Docket No. 2021–012). G.P.N. owns shares in Acrea 3D, a company commercializing microfluidic 3D printing.
Figures
Similar articles
-
Engineering de novo assembly of fetal pulmonary organoids.Tissue Eng Part A. 2014 Nov;20(21-22):2892-907. doi: 10.1089/ten.TEA.2014.0085. Epub 2014 Jun 25. Tissue Eng Part A. 2014. PMID: 24825442 Free PMC article.
-
Human airway organoid engineering as a step toward lung regeneration and disease modeling.Biomaterials. 2017 Jan;113:118-132. doi: 10.1016/j.biomaterials.2016.10.046. Epub 2016 Oct 28. Biomaterials. 2017. PMID: 27815996 Free PMC article.
-
Disease modeling of pulmonary fibrosis using human pluripotent stem cell-derived alveolar organoids.Stem Cell Reports. 2021 Dec 14;16(12):2973-2987. doi: 10.1016/j.stemcr.2021.10.015. Epub 2021 Nov 18. Stem Cell Reports. 2021. PMID: 34798066 Free PMC article.
-
Bioengineering tissue morphogenesis and function in human neural organoids.Semin Cell Dev Biol. 2021 Mar;111:52-59. doi: 10.1016/j.semcdb.2020.05.025. Epub 2020 Jun 12. Semin Cell Dev Biol. 2021. PMID: 32540123 Free PMC article. Review.
-
Extracellular matrix requirements for gastrointestinal organoid cultures.Biomaterials. 2021 Sep;276:121020. doi: 10.1016/j.biomaterials.2021.121020. Epub 2021 Jul 10. Biomaterials. 2021. PMID: 34280822 Review.
Cited by
-
Best Practices in the Development and Use of Experimental Models of Bacterial Pneumonia: An Official American Thoracic Society Workshop Report.Am J Respir Cell Mol Biol. 2025 Aug;73(2):178-199. doi: 10.1165/rcmb.2025-0322ST. Am J Respir Cell Mol Biol. 2025. PMID: 40748059 Free PMC article.
-
De novo design of highly selective miniprotein inhibitors of integrins αvβ6 and αvβ8.bioRxiv [Preprint]. 2023 Jun 12:2023.06.12.544624. doi: 10.1101/2023.06.12.544624. bioRxiv. 2023. Update in: Nat Commun. 2023 Sep 13;14(1):5660. doi: 10.1038/s41467-023-41272-z. PMID: 37398153 Free PMC article. Updated. Preprint.
-
CEBPA Overexpression Enhances β-Cell Proliferation and Survival.Biology (Basel). 2024 Feb 9;13(2):110. doi: 10.3390/biology13020110. Biology (Basel). 2024. PMID: 38392328 Free PMC article.
-
Potential long-term effects of SARS-CoV-2 infection on the pulmonary vasculature: Multilayered cross-talks in the setting of coinfections and comorbidities.PLoS Pathog. 2023 Jan 12;19(1):e1011063. doi: 10.1371/journal.ppat.1011063. eCollection 2023 Jan. PLoS Pathog. 2023. PMID: 36634048 Free PMC article. Review.
-
The evolution of in vitro models of lung fibrosis: promising prospects for drug discovery.Eur Respir Rev. 2024 Jan 17;33(171):230127. doi: 10.1183/16000617.0127-2023. Print 2024 Jan 31. Eur Respir Rev. 2024. PMID: 38232990 Free PMC article. Review.
References
-
- Rousselle P, Scoazec JY, Laminin 332 in cancer: when the extracellular matrix turns signals from cell anchorage to cell movement, Semin. Cancer Biol 62 (2020) 149–165. - PubMed
-
- Buchheit CL, Rayavarapu RR, Schafer ZT, The regulation of cancer cell death and metabolism by extracellular matrix attachment, Semin. Cell Dev. Biol 23 (4) (2012) 402–411. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
