

Skeletal muscle mitoribosomal defects are linked to low bone mass caused by bone marrow inflammation in male mice
- PMID: 35306755
- PMCID: PMC9178379
- DOI: 10.1002/jcsm.12975
Skeletal muscle mitoribosomal defects are linked to low bone mass caused by bone marrow inflammation in male mice
Abstract
Background: Mitochondrial oxidative phosphorylation (OxPhos) is a critical regulator of skeletal muscle mass and function. Although muscle atrophy due to mitochondrial dysfunction is closely associated with bone loss, the biological characteristics of the relationship between muscle and bone remain obscure. We showed that muscle atrophy caused by skeletal muscle-specific CR6-interacting factor 1 knockout (MKO) modulates the bone marrow (BM) inflammatory response, leading to low bone mass.
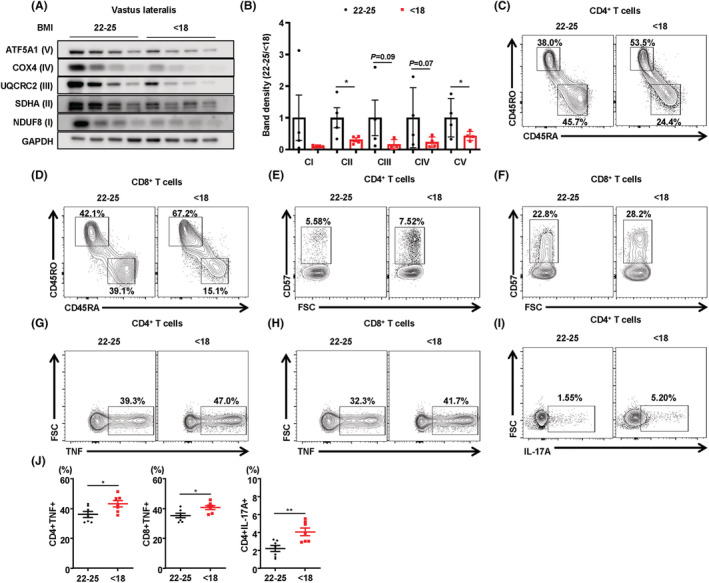
Methods: MKO mice with lower muscle OxPhos were fed a normal chow or high-fat diet and then evaluated for muscle mass and function, and bone mineral density. Immunophenotyping of BM immune cells was also performed. BM transcriptomic analysis was used to identify key factors regulating bone mass in MKO mice. To determine the effects of BM-derived CXCL12 (C-X-C motif chemokine ligand 12) on regulation of bone homeostasis, a variety of BM niche-resident cells were treated with recombinant CXCL12. Vastus lateralis muscle and BM immune cell samples from 14 patients with hip fracture were investigated to examine the association between muscle function and BM inflammation.
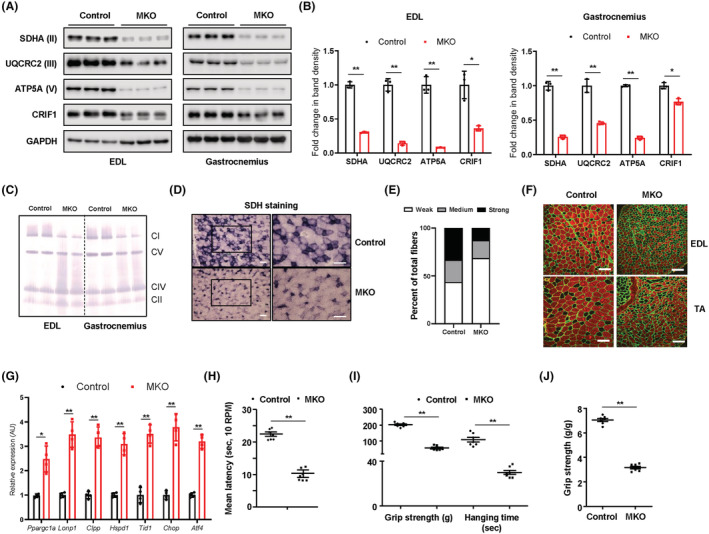
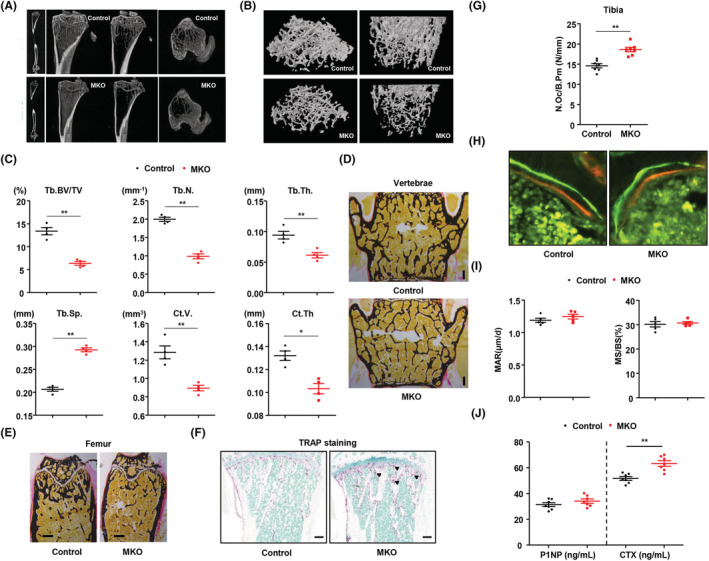
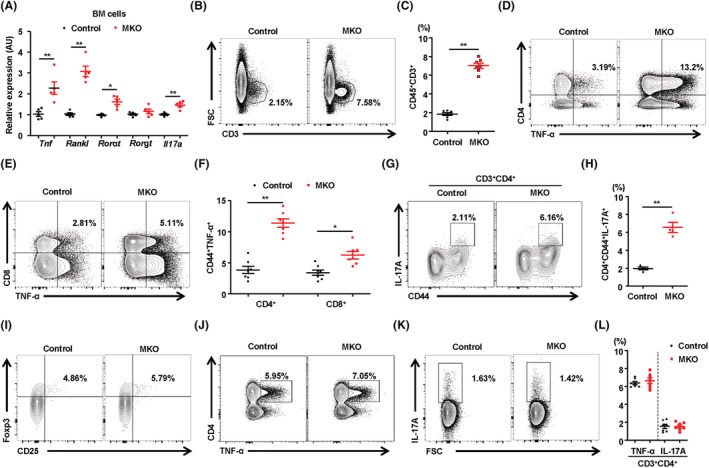
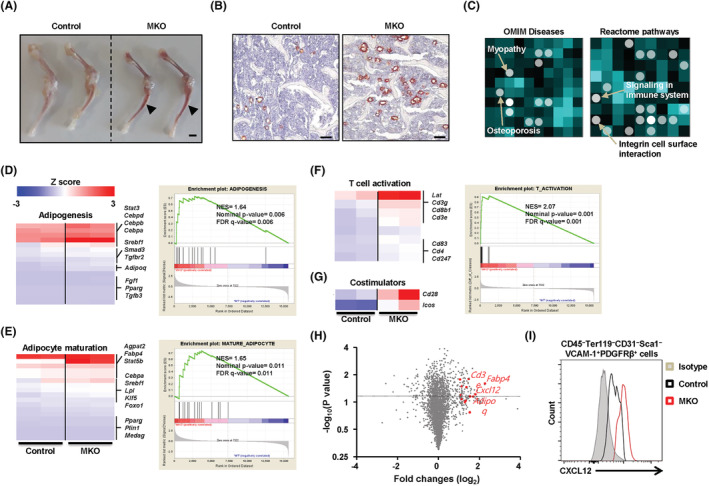
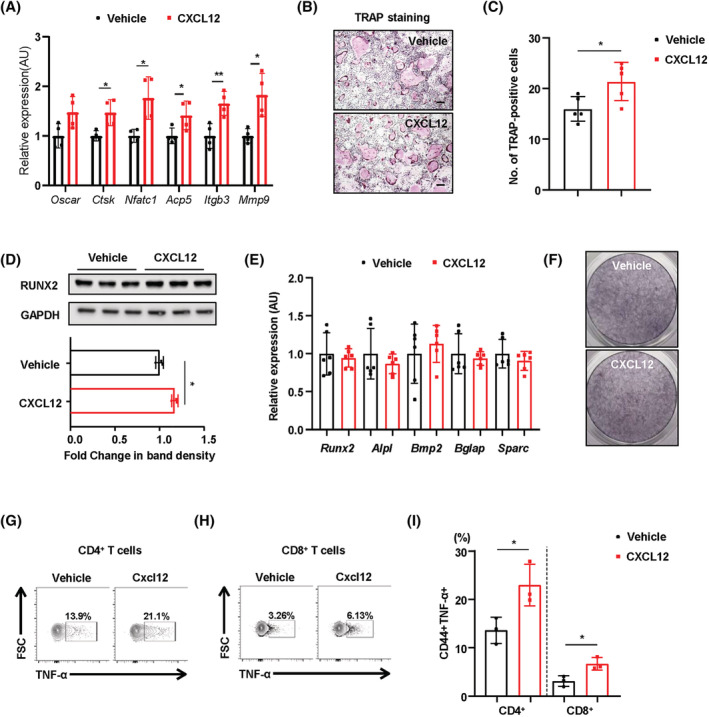
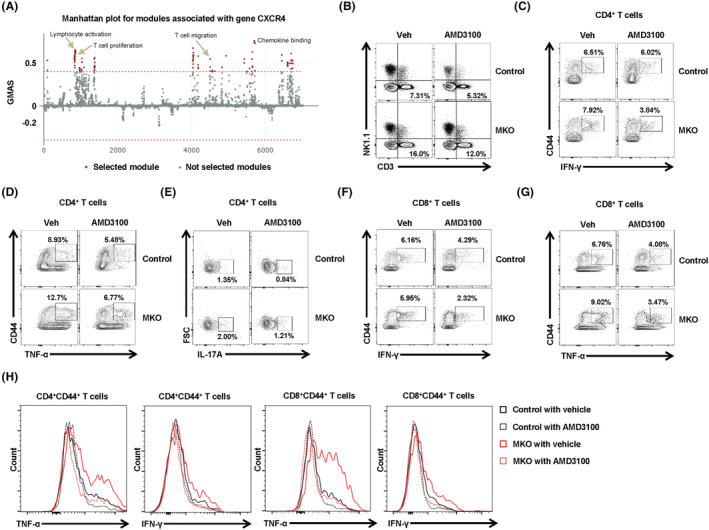
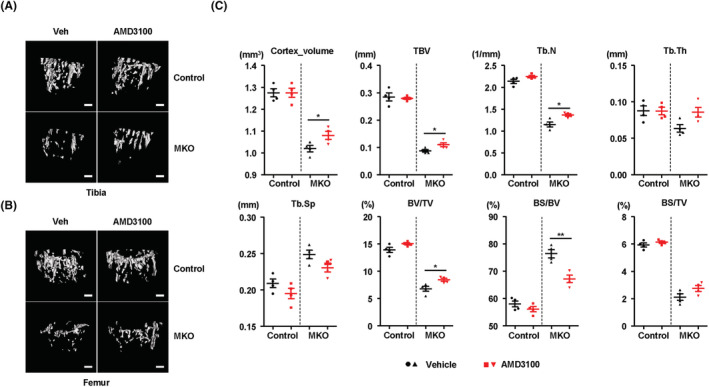
Results: MKO mice exhibited significant reductions in both muscle mass and expression of OxPhos subunits but increased transcription of mitochondrial stress response-related genes in the extensor digitorum longus (P < 0.01). MKO mice showed a decline in grip strength and a higher drop rate in the wire hanging test (P < 0.01). Micro-computed tomography and von Kossa staining revealed that MKO mice developed a low mass phenotype in cortical and trabecular bone (P < 0.01). Transcriptomic analysis of the BM revealed that mitochondrial stress responses in skeletal muscles induce an inflammatory response and adipogenesis in the BM and that the CXCL12-CXCR4 (C-X-C chemokine receptor 4) axis is important for T-cell homing to the BM. Antagonism of CXCR4 attenuated BM inflammation and increased bone mass in MKO mice. In humans, patients with low body mass index (BMI = 17.2 ± 0.42 kg/m2 ) harboured a larger population of proinflammatory and cytotoxic senescent T-cells in the BMI (P < 0.05) and showed reduced expression of OxPhos subunits in the vastus lateralis, compared with controls with a normal BMI (23.7 ± 0.88 kg/m2 ) (P < 0.01).
Conclusions: Defects in muscle mitochondrial OxPhos promote BM inflammation in mice, leading to decreased bone mass. Muscle mitochondrial dysfunction is linked to BM inflammatory cytokine secretion via the CXCL12-CXCR4 signalling axis, which is critical for inducing low bone mass.
Keywords: Bone loss; Bone marrow; Inflammation; Mitochondria.
© 2022 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Giresi PG, Stevenson EJ, Theilhaber J, Koncarevic A, Parkington J, Fielding RA, et al. Identification of a molecular signature of sarcopenia. Physiol Genomics 2005;21:253–263. - PubMed
-
- Verschueren S, Gielen E, O'Neill TW, Pye SR, Adams JE, Ward KA, et al. Sarcopenia and its relationship with bone mineral density in middle‐aged and elderly European men. Osteoporos Int 2013;24:87–98. - PubMed
-
- Trifunovic A, Wredenberg A, Falkenberg M, Spelbrink JN, Rovio AT, Bruder CE, et al. Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature 2004;429:417–423. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
