

How Does the Central Nervous System for Posture and Locomotion Cope With Damage-Induced Neural Asymmetry?
- PMID: 35308565
- PMCID: PMC8927091
- DOI: 10.3389/fnsys.2022.828532
How Does the Central Nervous System for Posture and Locomotion Cope With Damage-Induced Neural Asymmetry?
Abstract
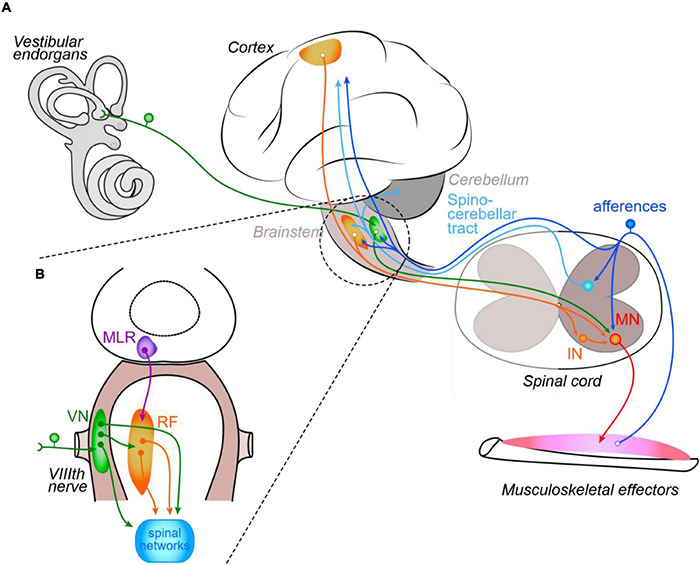
In most vertebrates, posture and locomotion are achieved by a biomechanical apparatus whose effectors are symmetrically positioned around the main body axis. Logically, motor commands to these effectors are intrinsically adapted to such anatomical symmetry, and the underlying sensory-motor neural networks are correspondingly arranged during central nervous system (CNS) development. However, many developmental and/or life accidents may alter such neural organization and acutely generate asymmetries in motor operation that are often at least partially compensated for over time. First, we briefly present the basic sensory-motor organization of posturo-locomotor networks in vertebrates. Next, we review some aspects of neural plasticity that is implemented in response to unilateral central injury or asymmetrical sensory deprivation in order to substantially restore symmetry in the control of posturo-locomotor functions. Data are finally discussed in the context of CNS structure-function relationship.
Keywords: development; injury; motor recovery; neuronal networks; sensory-motor integration.
Copyright © 2022 Le Ray and Guayasamin.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




References
-
- Alvarez F. J., Bullinger K. L., Titus H. E., Nardelli P., Cope T. C. (2010). Permanent reorganization of Ia afferent synapses on motoneurons after peripheral nerve injuries: monosynaptic stretch reflexes after nerve injuries. Ann. N. Y. Acad. Sci. 1198 231–241. 10.1111/j.1749-6632.2010.05459.x - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources