

Microtubule Anchoring: Attaching Dynamic Polymers to Cellular Structures
- PMID: 35309944
- PMCID: PMC8927778
- DOI: 10.3389/fcell.2022.867870
Microtubule Anchoring: Attaching Dynamic Polymers to Cellular Structures
Abstract
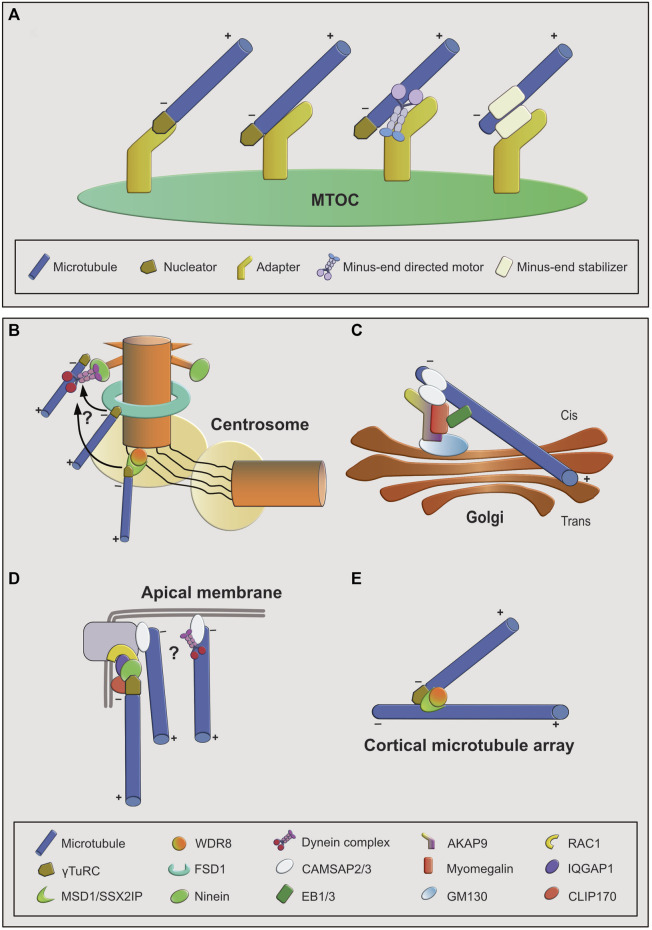
Microtubules are dynamic, filamentous polymers composed of α- and β-tubulin. Arrays of microtubules that have a specific polarity and distribution mediate essential processes such as intracellular transport and mitotic chromosome segregation. Microtubule arrays are generated with the help of microtubule organizing centers (MTOC). MTOCs typically combine two principal activities, the de novo formation of microtubules, termed nucleation, and the immobilization of one of the two ends of microtubules, termed anchoring. Nucleation is mediated by the γ-tubulin ring complex (γTuRC), which, in cooperation with its recruitment and activation factors, provides a template for α- and β-tubulin assembly, facilitating formation of microtubule polymer. In contrast, the molecules and mechanisms that anchor newly formed microtubules at MTOCs are less well characterized. Here we discuss the mechanistic challenges underlying microtubule anchoring, how this is linked with the molecular activities of known and proposed anchoring factors, and what consequences defective microtubule anchoring has at the cellular and organismal level.
Keywords: MTOC; anchoring; centrosome; microtubule; nucleation.
Copyright © 2022 Vineethakumari and Lüders.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources