

Helping behavior in prairie voles: A model of empathy and the importance of oxytocin
- PMID: 35310938
- PMCID: PMC8931361
- DOI: 10.1016/j.isci.2022.103991
Helping behavior in prairie voles: A model of empathy and the importance of oxytocin
Abstract
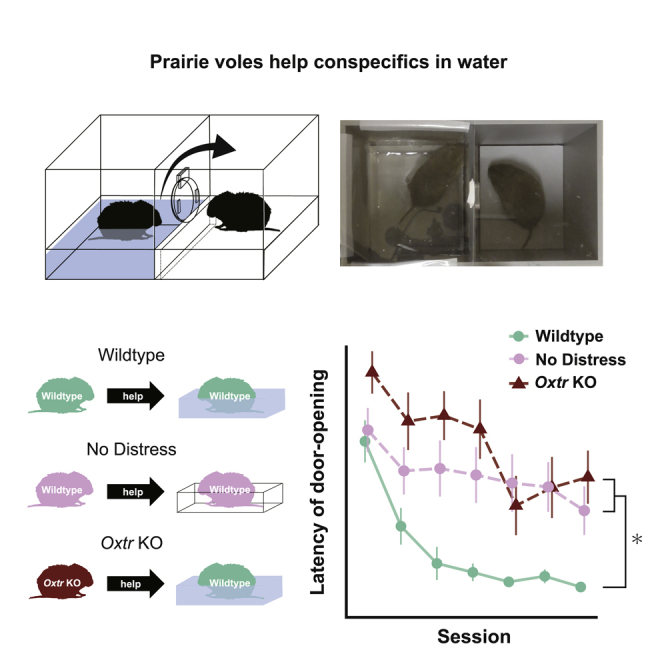
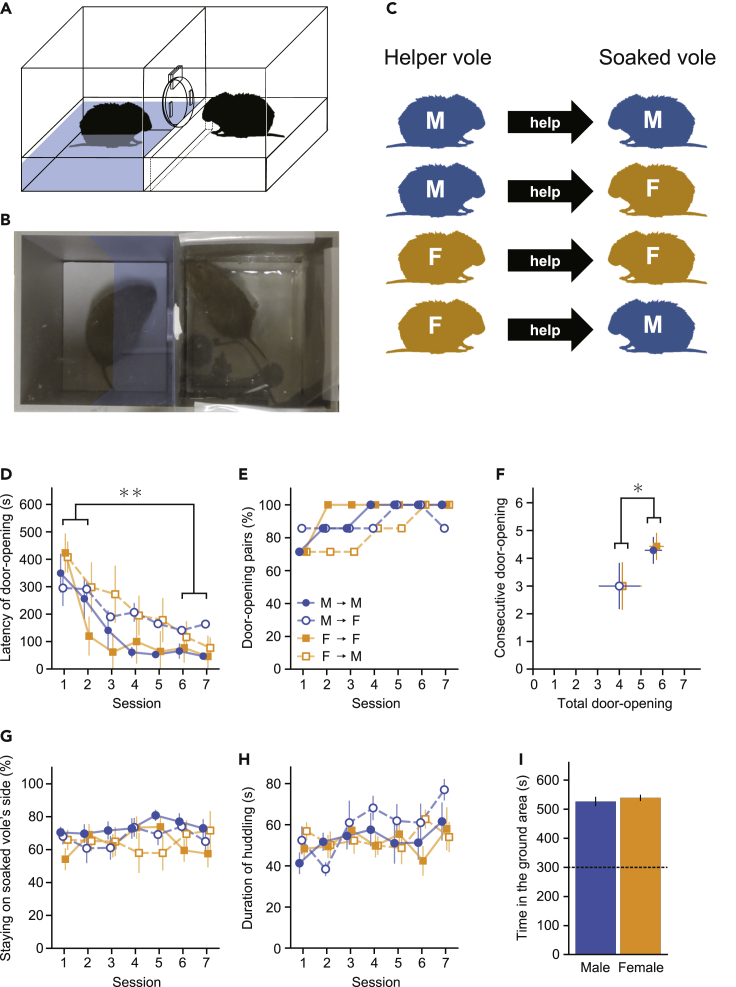
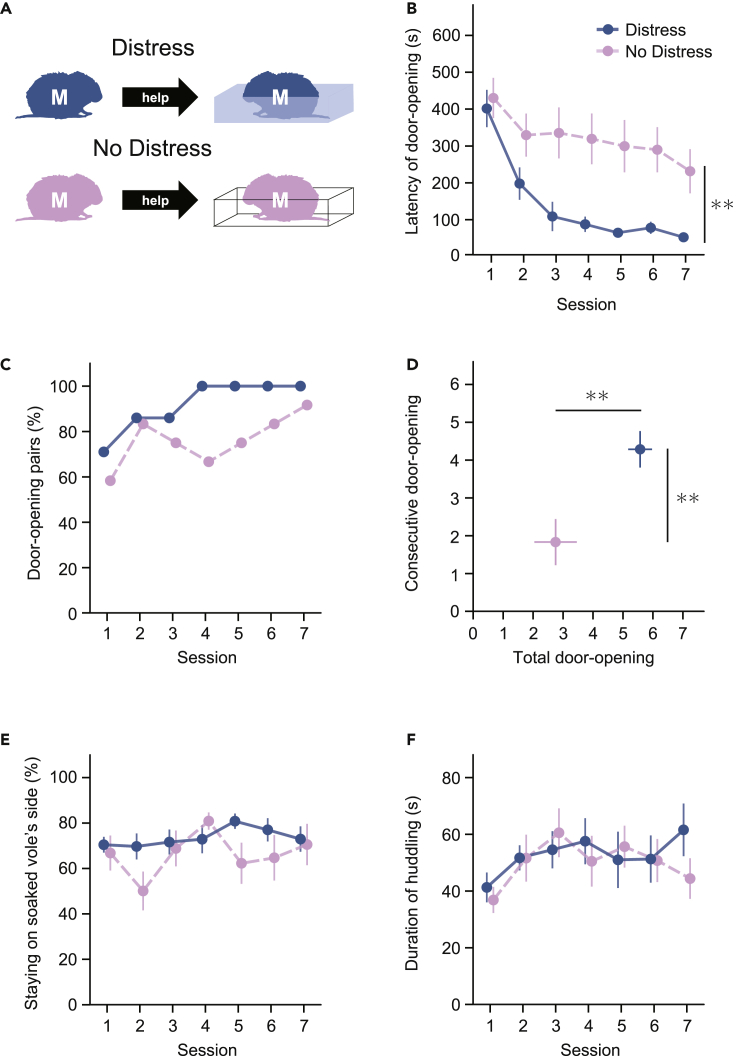
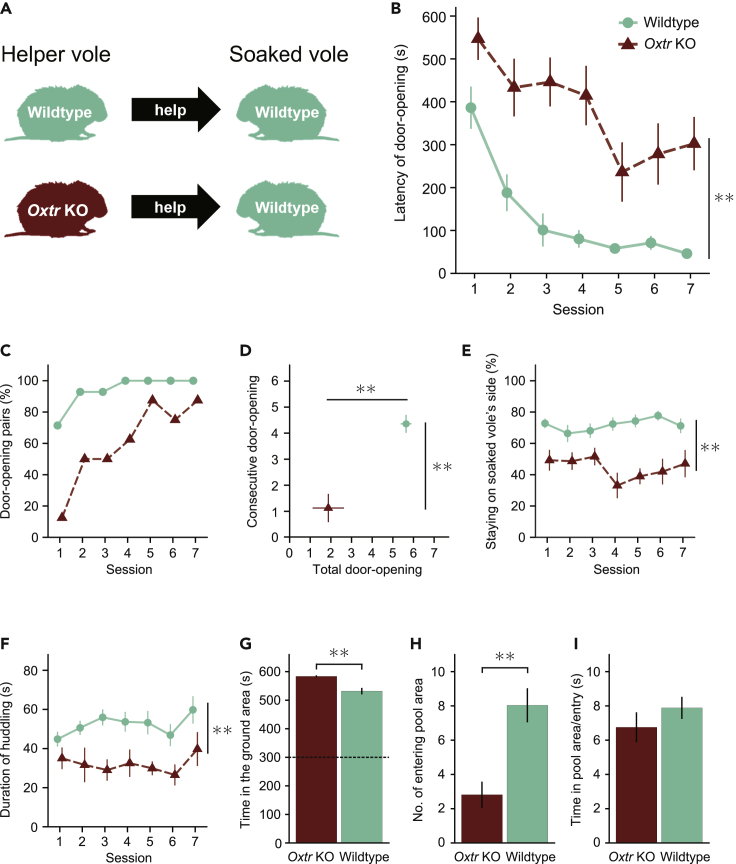
Several studies suggest that rodents show empathic responses and helping behavior toward others. We examined whether prairie voles would help conspecifics who were soaked in water by opening a door to a safe area. Door-opening latency decreased as task sessions progressed. Female and male voles stayed close to the soaked voles' side at equal rates and opened the door with similar latencies. When the conspecific was not soaked in water, the door-opening latency did not decrease. This suggests that the distress of the conspecific is necessary for learning to open the door and that the door-opening performed by prairie voles corresponds to helping behavior. Additionally, we examined the helping behavior in prairie voles in which oxytocin receptors were genetically knocked out. Oxytocin receptor knockout voles demonstrated less learning of the door-opening behavior and less interest in soaked conspecifics. This suggests that oxytocin is important for the emergence of helping behavior.
Keywords: Evolutionary biology; rodent behavior.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Rats demonstrate helping behavior toward a soaked conspecific.Anim Cogn. 2015 Sep;18(5):1039-47. doi: 10.1007/s10071-015-0872-2. Epub 2015 May 12. Anim Cogn. 2015. PMID: 25964095
-
Examining Empathy Through Consolation Behavior in Prairie Voles.J Undergrad Neurosci Educ. 2021 Jun 20;19(2):R35-R38. eCollection 2021 Spring. J Undergrad Neurosci Educ. 2021. PMID: 34552447 Free PMC article.
-
Oxytocin administration modulates rats' helping behavior depending on social context.Neurosci Res. 2020 Apr;153:56-61. doi: 10.1016/j.neures.2019.04.001. Epub 2019 Apr 3. Neurosci Res. 2020. PMID: 30953682
-
Neurochemical regulation of pair bonding in male prairie voles.Physiol Behav. 2004 Nov 15;83(2):319-28. doi: 10.1016/j.physbeh.2004.08.024. Physiol Behav. 2004. PMID: 15488548 Review.
-
Cellular mechanisms of social attachment.Horm Behav. 2001 Sep;40(2):133-8. doi: 10.1006/hbeh.2001.1691. Horm Behav. 2001. PMID: 11534973 Review.
Cited by
-
Third-party punishment-like behavior in a rat model.Sci Rep. 2024 Sep 27;14(1):22310. doi: 10.1038/s41598-024-71748-x. Sci Rep. 2024. PMID: 39333120 Free PMC article.
-
Oxytocin receptors are widely distributed in the prairie vole (Microtus ochrogaster) brain: Relation to social behavior, genetic polymorphisms, and the dopamine system.J Comp Neurol. 2022 Nov;530(16):2881-2900. doi: 10.1002/cne.25382. Epub 2022 Jun 28. J Comp Neurol. 2022. PMID: 35763609 Free PMC article.
-
Prosocial Helping Behavior: Conceptual Issues and Neural Mechanisms.Biol Psychiatry. 2025 May 15;97(10):961-970. doi: 10.1016/j.biopsych.2025.03.003. Epub 2025 Mar 14. Biol Psychiatry. 2025. PMID: 40090565 Review.
-
Oxytocin, Vasopressin, and Social Behavior: From Neural Circuits to Clinical Opportunities.Endocrinology. 2022 Sep 1;163(9):bqac111. doi: 10.1210/endocr/bqac111. Endocrinology. 2022. PMID: 35863332 Free PMC article. Review.
-
Why help others? Insights from rodent to human early childhood research.Front Behav Neurosci. 2023 Mar 21;17:1058352. doi: 10.3389/fnbeh.2023.1058352. eCollection 2023. Front Behav Neurosci. 2023. PMID: 37025110 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
