

Resistance-Guided Mining of Bacterial Genotoxins Defines a Family of DNA Glycosylases
- PMID: 35311535
- PMCID: PMC9040887
- DOI: 10.1128/mbio.03297-21
Resistance-Guided Mining of Bacterial Genotoxins Defines a Family of DNA Glycosylases
Abstract
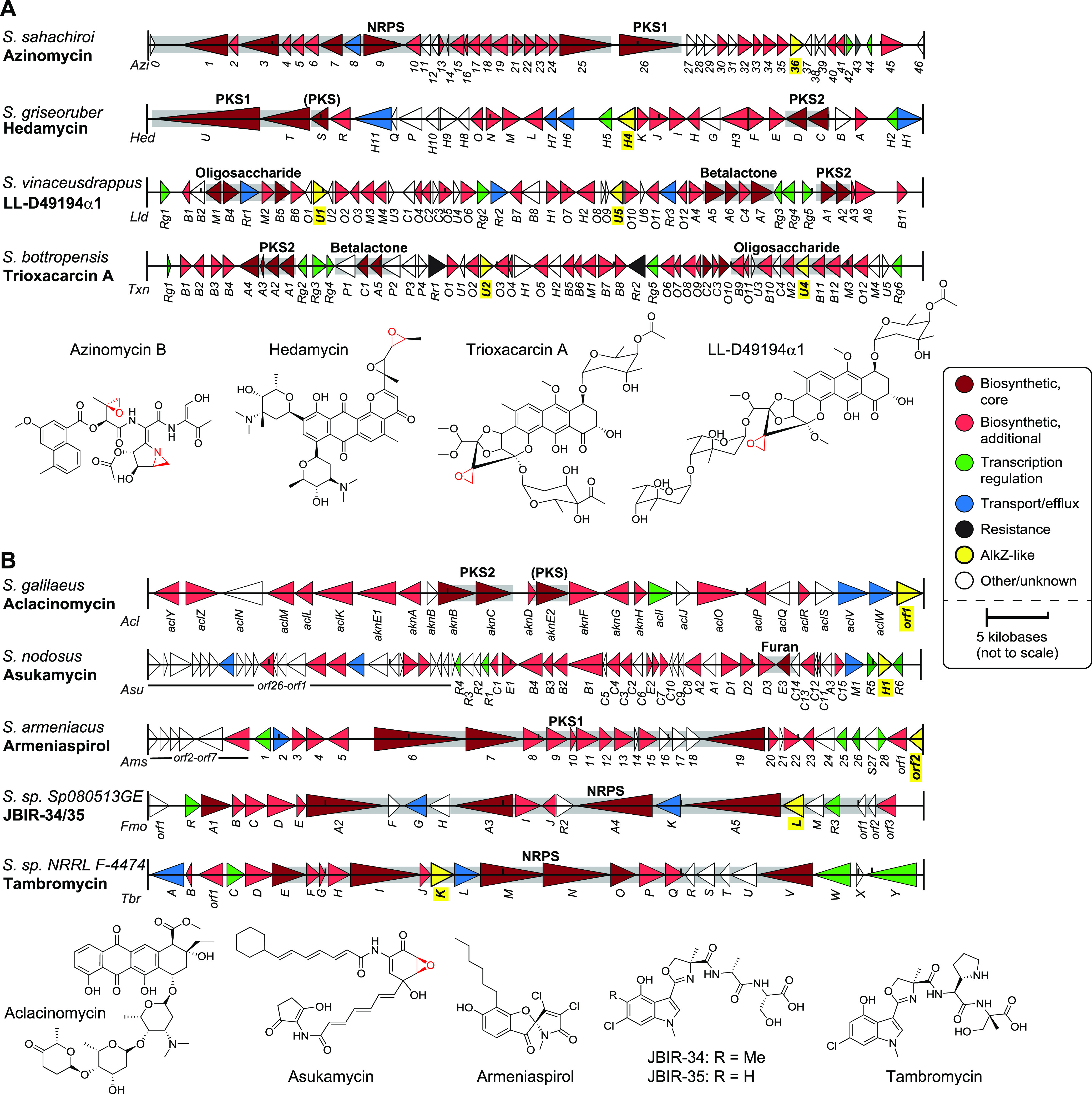
Unique DNA repair enzymes that provide self-resistance against therapeutically important, genotoxic natural products have been discovered in bacterial biosynthetic gene clusters (BGCs). Among these, the DNA glycosylase AlkZ is essential for azinomycin B production and belongs to the HTH_42 superfamily of uncharacterized proteins. Despite their widespread existence in antibiotic producers and pathogens, the roles of these proteins in production of other natural products are unknown. Here, we determine the evolutionary relationship and genomic distribution of all HTH_42 proteins from Streptomyces and use a resistance-based genome mining approach to identify homologs associated with known and uncharacterized BGCs. We find that AlkZ-like (AZL) proteins constitute one distinct HTH_42 subfamily and are highly enriched in BGCs and variable in sequence, suggesting each has evolved to protect against a specific secondary metabolite. As a validation of the approach, we show that the AZL protein, HedH4, associated with biosynthesis of the alkylating agent hedamycin, excises hedamycin-DNA adducts with exquisite specificity and provides resistance to the natural product in cells. We also identify a second, phylogenetically and functionally distinct subfamily whose proteins are never associated with BGCs, are highly conserved with respect to sequence and genomic neighborhood, and repair DNA lesions not associated with a particular natural product. This work delineates two related families of DNA repair enzymes-one specific for complex alkyl-DNA lesions and involved in self-resistance to antimicrobials and the other likely involved in protection against an array of genotoxins-and provides a framework for targeted discovery of new genotoxic compounds with therapeutic potential. IMPORTANCE Bacteria are rich sources of secondary metabolites that include DNA-damaging genotoxins with antitumor/antibiotic properties. Although Streptomyces produce a diverse number of therapeutic genotoxins, efforts toward targeted discovery of biosynthetic gene clusters (BGCs) producing DNA-damaging agents is lacking. Moreover, work on toxin-resistance genes has lagged behind our understanding of those involved in natural product synthesis. Here, we identified over 70 uncharacterized BGCs producing potentially novel genotoxins through resistance-based genome mining using the azinomycin B-resistance DNA glycosylase AlkZ. We validate our analysis by characterizing the enzymatic activity and cellular resistance of one AlkZ ortholog in the BGC of hedamycin, a potent DNA alkylating agent. Moreover, we uncover a second, phylogenetically distinct family of proteins related to Escherichia coli YcaQ, a DNA glycosylase capable of unhooking interstrand DNA cross-links, which differs from the AlkZ-like family in sequence, genomic location, proximity to BGCs, and substrate specificity. This work defines two families of DNA glycosylase for specialized repair of complex genotoxic natural products and generalized repair of a broad range of alkyl-DNA adducts and provides a framework for targeted discovery of new compounds with therapeutic potential.
Keywords: AlkZ; DNA cross-link; DNA glycosylase; DNA repair; HTH_42; Streptomyces; biosynthetic gene cluster; genotoxin; intecalator; natural product; phylogenetic tree; secondary metabolism; self-resistance.
Conflict of interest statement
The authors declare a conflict of interest. A.R. is a scientific consultant for LifeMine Therapeutics, Inc.
Figures




Similar articles
-
Structural evolution of a DNA repair self-resistance mechanism targeting genotoxic secondary metabolites.Nat Commun. 2021 Nov 26;12(1):6942. doi: 10.1038/s41467-021-27284-7. Nat Commun. 2021. PMID: 34836957 Free PMC article.
-
Structure of a DNA glycosylase that unhooks interstrand cross-links.Proc Natl Acad Sci U S A. 2017 Apr 25;114(17):4400-4405. doi: 10.1073/pnas.1703066114. Epub 2017 Apr 10. Proc Natl Acad Sci U S A. 2017. PMID: 28396405 Free PMC article.
-
Characterization of a novel DNA glycosylase from S. sahachiroi involved in the reduction and repair of azinomycin B induced DNA damage.Nucleic Acids Res. 2016 Jan 8;44(1):187-97. doi: 10.1093/nar/gkv949. Epub 2015 Sep 22. Nucleic Acids Res. 2016. PMID: 26400161 Free PMC article.
-
Genome mining of Streptomyces ambofaciens.J Ind Microbiol Biotechnol. 2014 Feb;41(2):251-63. doi: 10.1007/s10295-013-1379-y. Epub 2013 Nov 21. J Ind Microbiol Biotechnol. 2014. PMID: 24258629 Review.
-
Antibiotic export: transporters involved in the final step of natural product production.Microbiology (Reading). 2019 Aug;165(8):805-818. doi: 10.1099/mic.0.000794. Epub 2019 Apr 9. Microbiology (Reading). 2019. PMID: 30964430 Review.
Cited by
-
Biosynthesis of DNA-Alkylating Antitumor Natural Products.Molecules. 2022 Sep 27;27(19):6387. doi: 10.3390/molecules27196387. Molecules. 2022. PMID: 36234921 Free PMC article. Review.
-
Yatakemycin biosynthesis requires two deoxyribonucleases for toxin self-resistance.RSC Chem Biol. 2024 Dec 3;6(1):94-105. doi: 10.1039/d4cb00203b. eCollection 2025 Jan 2. RSC Chem Biol. 2024. PMID: 39649339 Free PMC article.
-
An interstrand DNA crosslink glycosylase aids Acinetobacter baumannii pathogenesis.Proc Natl Acad Sci U S A. 2024 Jul 2;121(27):e2402422121. doi: 10.1073/pnas.2402422121. Epub 2024 Jun 26. Proc Natl Acad Sci U S A. 2024. PMID: 38923984 Free PMC article.
-
Identification of the lydiamycin biosynthetic gene cluster in a plant pathogen guides structural revision and identification of molecular target.Proc Natl Acad Sci U S A. 2025 May 27;122(21):e2424388122. doi: 10.1073/pnas.2424388122. Epub 2025 May 19. Proc Natl Acad Sci U S A. 2025. PMID: 40388608 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
