

A pan-genotype hepatitis C virus viral vector vaccine generates T cells and neutralizing antibodies in mice
- PMID: 35313015
- PMCID: PMC9790311
- DOI: 10.1002/hep.32470
A pan-genotype hepatitis C virus viral vector vaccine generates T cells and neutralizing antibodies in mice
Abstract
Background and aims: A prophylactic vaccine targeting multiple HCV genotypes (gt) is urgently required to meet World Health Organization elimination targets. Neutralizing antibodies (nAbs) and CD4+ and CD8+ T cells are associated with spontaneous clearance of HCV, and each may contribute to protective immunity. However, current vaccine candidates generate either nAbs or T cells targeting genetically variable epitopes and have failed to show efficacy in human trials. We have previously shown that a simian adenovirus vector (ChAdOx1) encoding conserved sequences across gt1-6 (ChAd-Gt1-6), and separately gt-1a E2 protein with variable regions deleted (E2Δ123HMW ), generates pan-genotypic T cells and nAbs, respectively. We now aim to develop a vaccine to generate both viral-specific B- and T-cell responses concurrently.
Approach and results: We show that vaccinating with ChAd-Gt1-6 and E2Δ123HMW sequentially in mice generates T-cell and antibody (Ab) responses comparable to either vaccine given alone. We encoded E2Δ123 in ChAdOx1 (ChAd-E2Δ123) and show that this, given with an E2Δ123HMW protein boost, induces greater CD81-E2 inhibitory and HCV-pseudoparticle nAb titers compared to the E2Δ123HMW prime boost. We developed bivalent viral vector vaccines (ChAdOx1 and modified vaccinia Ankara [MVA]) encoding both Gt1-6 and E2Δ123 immunogens (Gt1-6-E2Δ123) generating polyfunctional CD4+ and CD8+ T cells and nAb titers in prime/boost strategies. This approach generated nAb responses comparable to monovalent E2Δ123 ChAd/MVA vaccines and superior to three doses of recombinant E2Δ123HMW protein, while also generating high-magnitude T-cell responses.
Conclusions: These data are an important step forward for the development of a pan-genotype HCV vaccine to elicit T cells and nAbs for future assessment in humans.
© 2022 The Authors. Hepatology published by Wiley Periodicals LLC on behalf of American Association for the Study of Liver Diseases.
Conflict of interest statement
T.D., S.C., and E.B. are all contributors or inventors on patent PCT/GB2017/050840 that describes the conserved segment HCV Gt1‐6 T‐cell vaccine used in this study. H.D. and P.P. are named inventors on patents PCT/AU2007/001221 and PCT/AU2011/001534 that describe E2 antigens used in this study. H.D., E.B., and S.C. are named inventors of PCT/AU2021/050437.
Figures
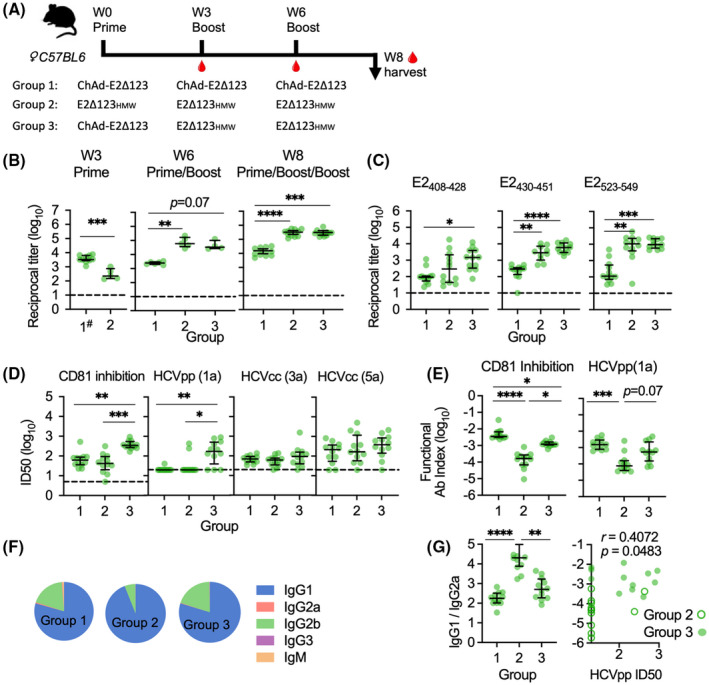
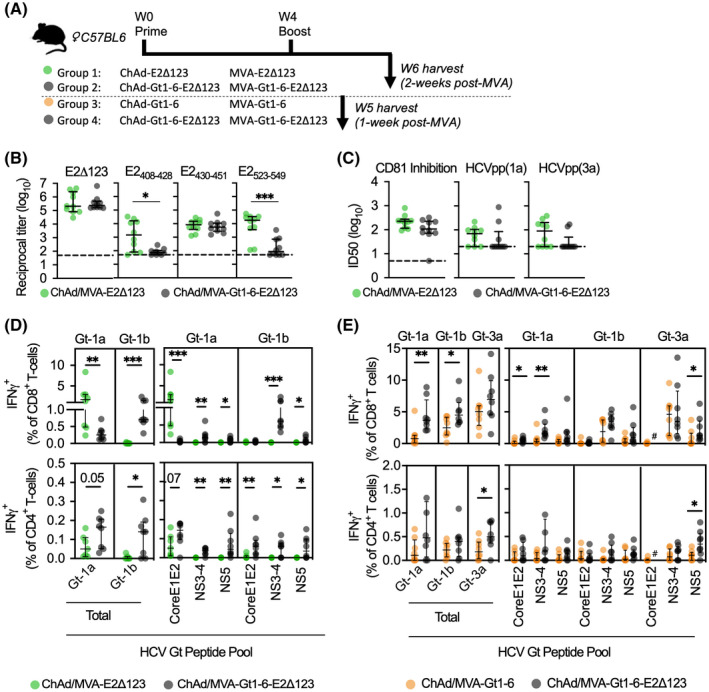
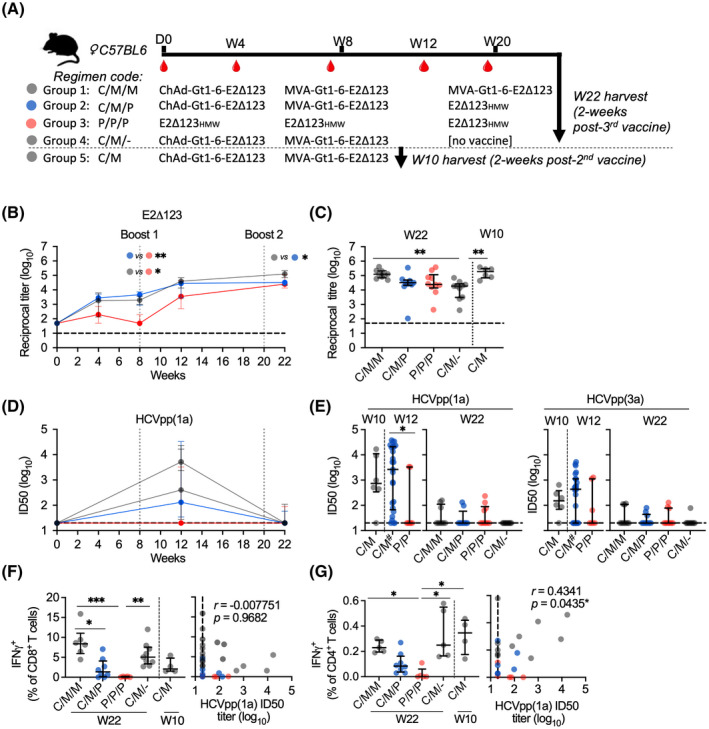
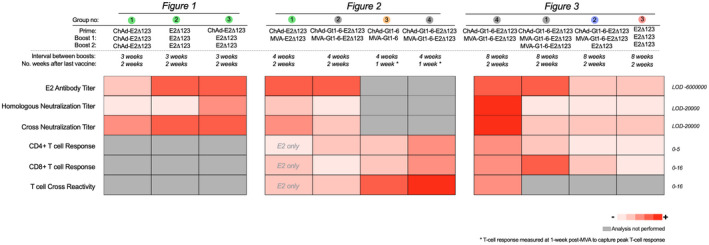
Comment in
-
Another important step toward a prophylactic vaccine against hepatitis C.Hepatology. 2022 Oct;76(4):917-919. doi: 10.1002/hep.32557. Epub 2022 May 19. Hepatology. 2022. PMID: 35503709 No abstract available.
References
-
- World Health Organization . Global Hepatitis Report 2017. Geneva, Switzerland: World Health Organization; 2017.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
