

Probing single molecule mechanical interactions of syntaxin 1A with native synaptobrevin 2 residing on a secretory vesicle
- PMID: 35314381
- PMCID: PMC9119915
- DOI: 10.1016/j.ceca.2022.102570
Probing single molecule mechanical interactions of syntaxin 1A with native synaptobrevin 2 residing on a secretory vesicle
Abstract
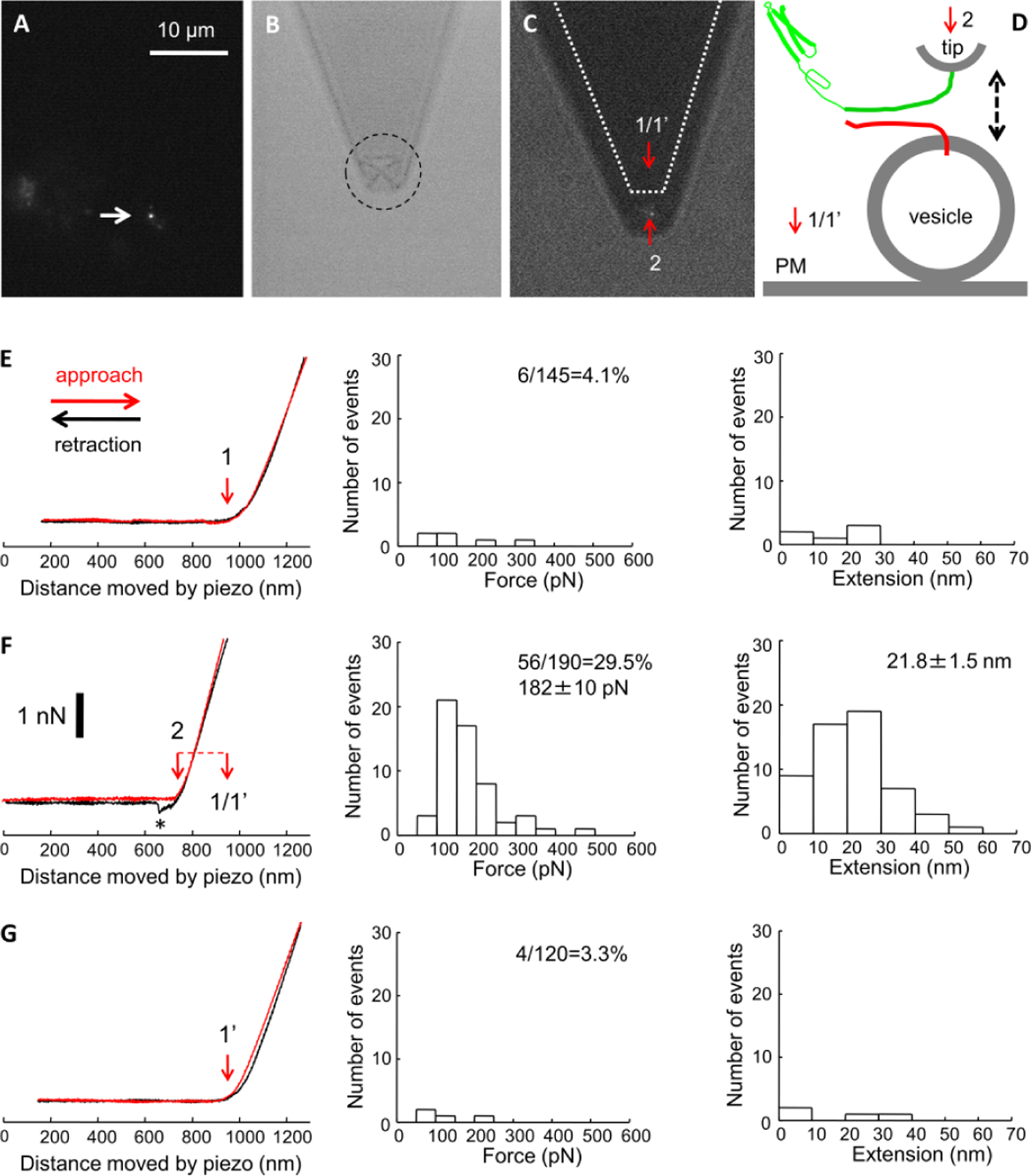
Interactive mechanical forces between pairs of individual SNARE proteins synaptobrevin 2 (Sb2) and syntaxin 1A (Sx1A) may be sufficient to mediate vesicle docking. This notion, based on force spectroscopy single molecule measurements probing recombinant Sx1A an Sb2 in silico, questioned a predominant view of docking via the ternary SNARE complex formation, which includes an assembly of the intermediate cis binary complex between Sx1A and SNAP25 on the plasma membrane to engage Sb2 on the vesicle. However, whether a trans binary Sx1A-Sb2 complex alone could mediate vesicle docking in a cellular environment remains unclear. To address this issue, we used atomic force microscopy (AFM) in the force spectroscopy mode combined with fluorescence imaging. Using AFM tips functionalized with the full Sx1A cytosolic domain, we probed native Sb2 studding the membrane of secretory vesicles docked at the plasma membrane patches, referred to as "inside-out lawns", identified based on fluorescence stains and prepared from primary culture of lactotrophs. We recorded single molecule Sx1A-Sb2 mechanical interactions and obtained measurements of force (∼183 pN) and extension (∼21.6 nm) necessary to take apart Sx1A-Sb2 binding interactions formed at tip-vesicle contact. Measured interactive force between a single pair of Sx1A-Sb2 molecules is sufficient to hold a single secretory vesicle docked at the plasma membrane within distances up to that of the measured extension. This finding further advances a notion that native vesicle docking can be mediated by a single trans binary Sx1A-Sb2 complex in the absence of SNAP25.
Keywords: Atomic force microscopy; Docking; Exocytosis; Force spectroscopy; Membrane lawns; Nanomanipulation; SNARE proteins; Single molecule measurements; Vesicle docking.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflicts of interest
All authors declare no conflicts of interest.
Figures


Similar articles
-
Single molecule mechanical probing of the SNARE protein interactions.Biophys J. 2006 Jul 15;91(2):744-58. doi: 10.1529/biophysj.105.073312. Epub 2006 Apr 28. Biophys J. 2006. PMID: 16648158 Free PMC article.
-
Single-molecule measurements of dissociation rates and energy landscapes of binary trans snare complexes in parallel versus antiparallel orientation.Biophys J. 2011 Oct 19;101(8):1854-62. doi: 10.1016/j.bpj.2011.08.019. Biophys J. 2011. PMID: 22004738 Free PMC article.
-
Ternary SNARE complexes in parallel versus anti-parallel orientation: examination of their disassembly using single-molecule force spectroscopy.Cell Calcium. 2012 Sep-Oct;52(3-4):241-9. doi: 10.1016/j.ceca.2012.03.008. Epub 2012 Apr 21. Cell Calcium. 2012. PMID: 22525946
-
Core proteins of the secretory machinery.Handb Exp Pharmacol. 2008;(184):107-27. doi: 10.1007/978-3-540-74805-2_5. Handb Exp Pharmacol. 2008. PMID: 18064413 Review.
-
Functional regulation of syntaxin-1: An underlying mechanism mediating exocytosis in neuroendocrine cells.Front Endocrinol (Lausanne). 2023 Jan 19;14:1096365. doi: 10.3389/fendo.2023.1096365. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36742381 Free PMC article. Review.
Cited by
-
Striatal Syntaxin 1A Is Associated with Development of Tourette Syndrome in an Iminodipropionitrile-Induced Animal Model.Dis Markers. 2022 Sep 14;2022:1148191. doi: 10.1155/2022/1148191. eCollection 2022. Dis Markers. 2022. PMID: 36157213 Free PMC article.
-
Forty years of the adrenal chromaffin cell through ISCCB meetings around the world.Pflugers Arch. 2023 Jun;475(6):667-690. doi: 10.1007/s00424-023-02793-0. Epub 2023 Mar 8. Pflugers Arch. 2023. PMID: 36884064 Free PMC article. Review.
References
-
- Sollner T, Whiteheart SW, Brunner M, Erdjument-Bromage H, Geromanos S, Tempst P, Rothman JE, SNAP receptors implicated in vesicle targeting and fusion, Nature, 362 (1993) 318–324. - PubMed
-
- Jahn R, Scheller RH, SNAREs--engines for membrane fusion, Nat Rev Mol Cell Biol, 7 (2006) 631–643. - PubMed
-
- Sutton RB, Fasshauer D, Jahn R, Brunger AT, Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 A resolution, Nature, 395 (1998) 347–353. - PubMed
-
- McNally JM, Woodbury DJ, Lemos JR, Syntaxin 1A Drives Fusion of Large Dense-Core Neurosecretory Granules Into a Planar Lipid Bilayer, Cell Biochem Biophys, 41 (2004) 11–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
