

A non-dividing cell population with high pyruvate dehydrogenase kinase activity regulates metabolic heterogeneity and tumorigenesis in the intestine
- PMID: 35314684
- PMCID: PMC8938512
- DOI: 10.1038/s41467-022-29085-y
A non-dividing cell population with high pyruvate dehydrogenase kinase activity regulates metabolic heterogeneity and tumorigenesis in the intestine
Abstract
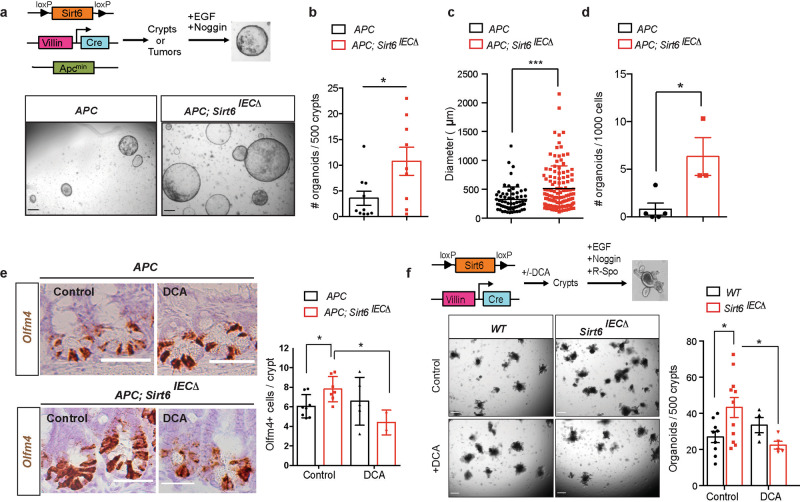
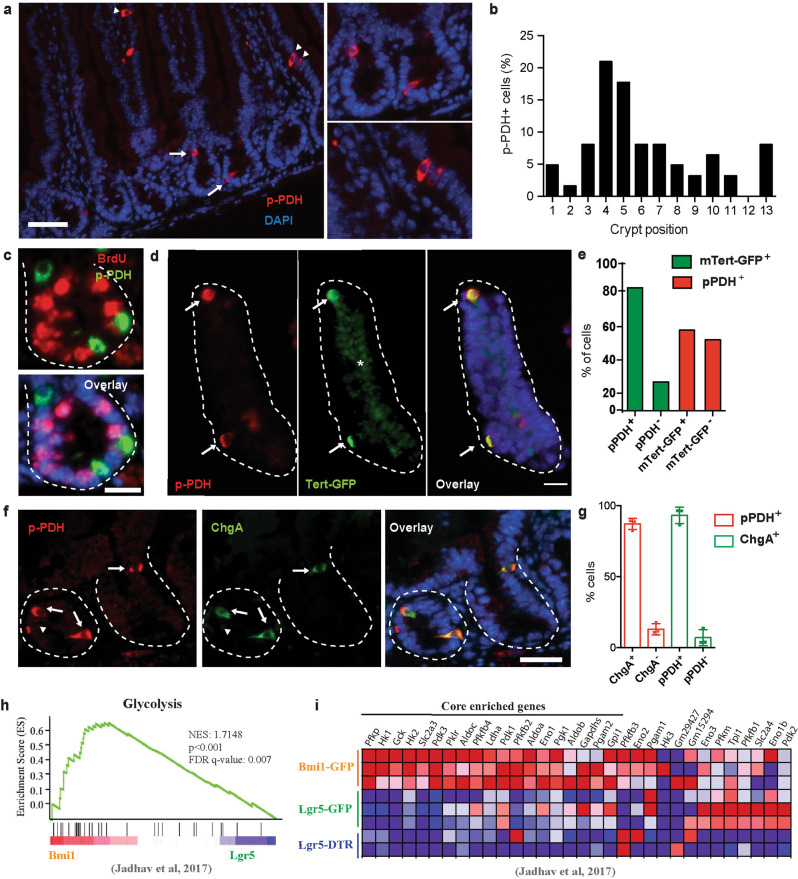
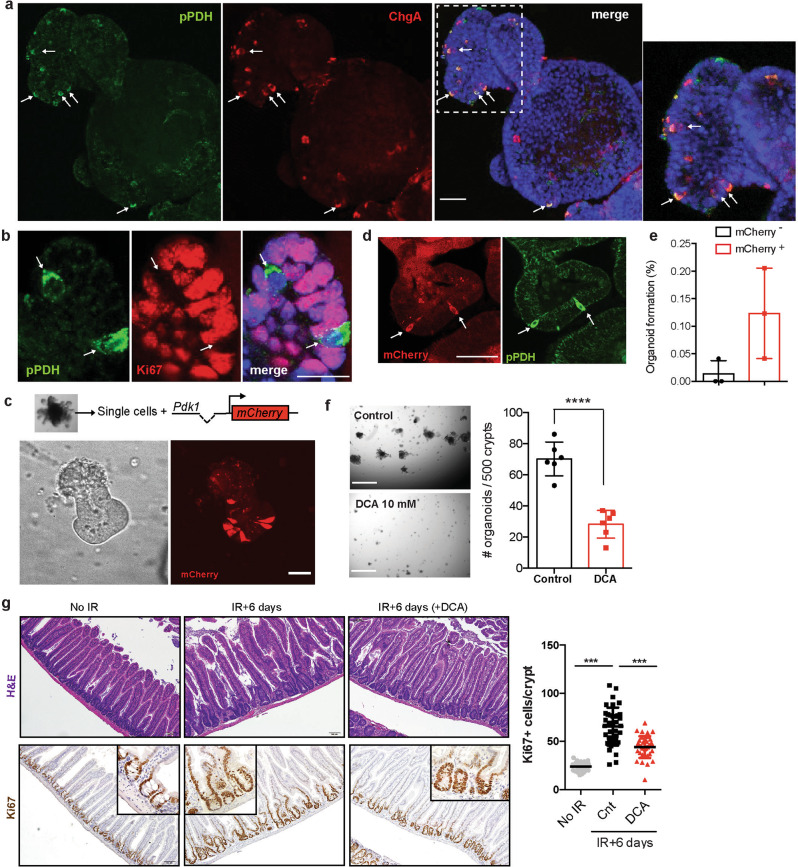
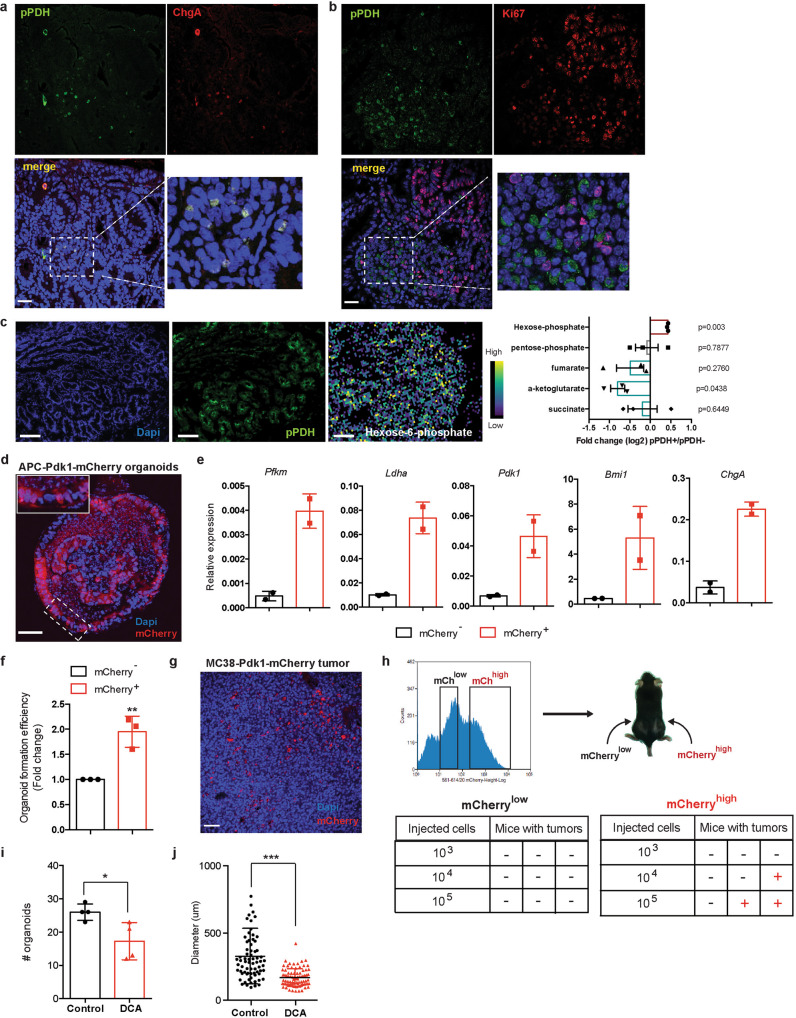
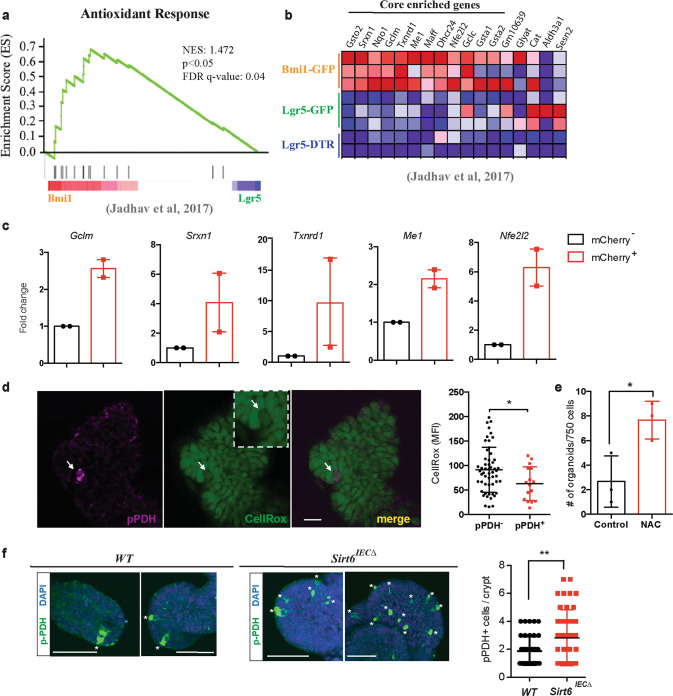
Although reprogramming of cellular metabolism is a hallmark of cancer, little is known about how metabolic reprogramming contributes to early stages of transformation. Here, we show that the histone deacetylase SIRT6 regulates tumor initiation during intestinal cancer by controlling glucose metabolism. Loss of SIRT6 results in an increase in the number of intestinal stem cells (ISCs), which translates into enhanced tumor initiating potential in APCmin mice. By tracking down the connection between glucose metabolism and tumor initiation, we find a metabolic compartmentalization within the intestinal epithelium and adenomas, where a rare population of cells exhibit features of Warburg-like metabolism characterized by high pyruvate dehydrogenase kinase (PDK) activity. Our results show that these cells are quiescent cells expressing +4 ISCs and enteroendocrine markers. Active glycolysis in these cells suppresses ROS accumulation and enhances their stem cell and tumorigenic potential. Our studies reveal that aerobic glycolysis represents a heterogeneous feature of cancer, and indicate that this metabolic adaptation can occur in non-dividing cells, suggesting a role for the Warburg effect beyond biomass production in tumors.
© 2022. The Author(s).
Conflict of interest statement
N.Y.R.A. is key opinion leader for Bruker Daltonics, scientific advisor to Invicro, and receives support from Thermo Finnegan and EMD Serono. R.M. is a founder of Galilei Biosciences.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 EB025823/EB/NIBIB NIH HHS/United States
- P30 DK040561/DK/NIDDK NIH HHS/United States
- R21 CA120342/CA/NCI NIH HHS/United States
- R01 CA235412/CA/NCI NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- R01 DK119488/DK/NIDDK NIH HHS/United States
- R01 CA175727/CA/NCI NIH HHS/United States
- P30 DK034854/DK/NIDDK NIH HHS/United States
- K99 CA252600/CA/NCI NIH HHS/United States
- P41 EB028741/EB/NIBIB NIH HHS/United States
- R01 GM128448/GM/NIGMS NIH HHS/United States
- U54 CA210180/CA/NCI NIH HHS/United States
- R00 CA252600/CA/NCI NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
