

Embryonic muscle splitting patterns reveal homologies of amniote forelimb muscles
- PMID: 35314784
- PMCID: PMC9090950
- DOI: 10.1038/s41559-022-01699-x
Embryonic muscle splitting patterns reveal homologies of amniote forelimb muscles
Abstract
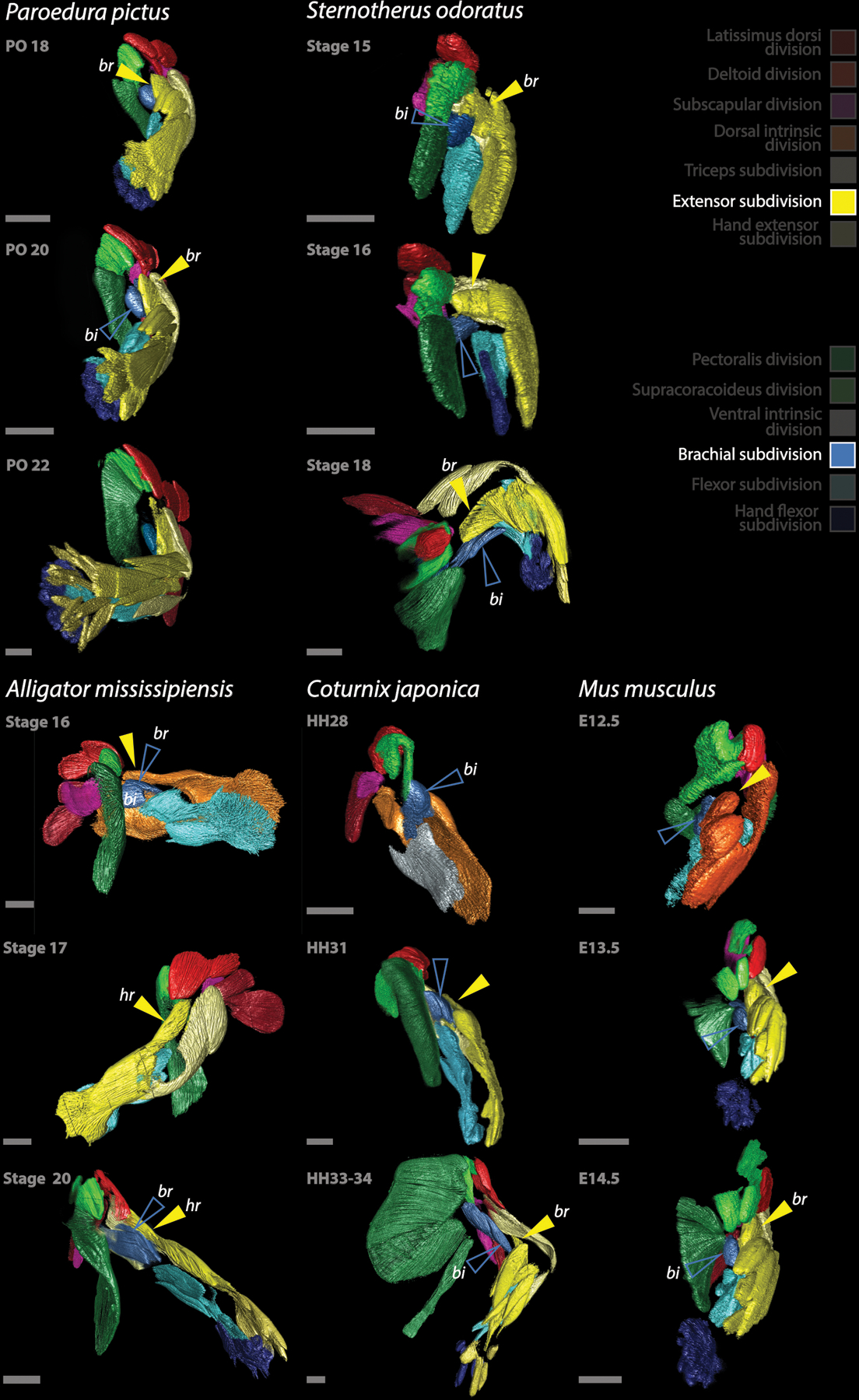
Limb muscles are remarkably complex and evolutionarily labile. Although their anatomy is of great interest for studies of the evolution of form and function, their homologies among major amniote clades have remained obscure. Studies of adult musculature are inconclusive owing to the highly derived morphology of modern amniote limbs but correspondences become increasingly evident earlier in ontogeny. We followed the embryonic development of forelimb musculature in representatives of six major amniote clades and found, contrary to current consensus, that these early splitting patterns are highly conserved across Amniota. Muscle mass cleavage patterns and topology are highly conserved in reptiles including birds, irrespective of their skeletal modifications: the avian flight apparatus results from slight early topological modifications that are exaggerated during ontogeny. Therian mammals, while conservative in their cleavage patterns, depart drastically from the ancestral amniote musculoskeletal organization in terms of topology. These topological changes occur through extension, translocation and displacement of muscle groups later in development. Overall, the simplicity underlying the apparent complexity of forelimb muscle development allows us to resolve conflicting hypotheses about homology and to trace the history of each individual forelimb muscle throughout the amniote radiations.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Conflict of interest
The authors declare no conflict of interest.
Figures














References
-
- Howell AB Morphogenesis of the shoulder architecture. Part IV. Reptilia. The Quarterly Review of Biology 11, 183–208 (1936).
-
- Howell AB Phylogeny of the distal musculature of the pectoral appendage. Journal of Morphology 60, 287–315 (1936).
-
- Howell AB Morphogenesis of the shoulder architecture: Aves. The Auk 54, 364–375 (1937).
-
- Howell AB Morphogenesis of the shoulder architecture. Part VI. Therian Mammalia. The Quarterly Review of Biology 12, 440–463 (1937).
-
- Howell AB Morphogenesis of the shoulder architecture. Part V. Monotremata. The Quarterly Review of Biology 12, 191–205 (1937).
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
