

Phylogenomic analyses of echinoid diversification prompt a re-evaluation of their fossil record
- PMID: 35315317
- PMCID: PMC8940180
- DOI: 10.7554/eLife.72460
Phylogenomic analyses of echinoid diversification prompt a re-evaluation of their fossil record
Abstract
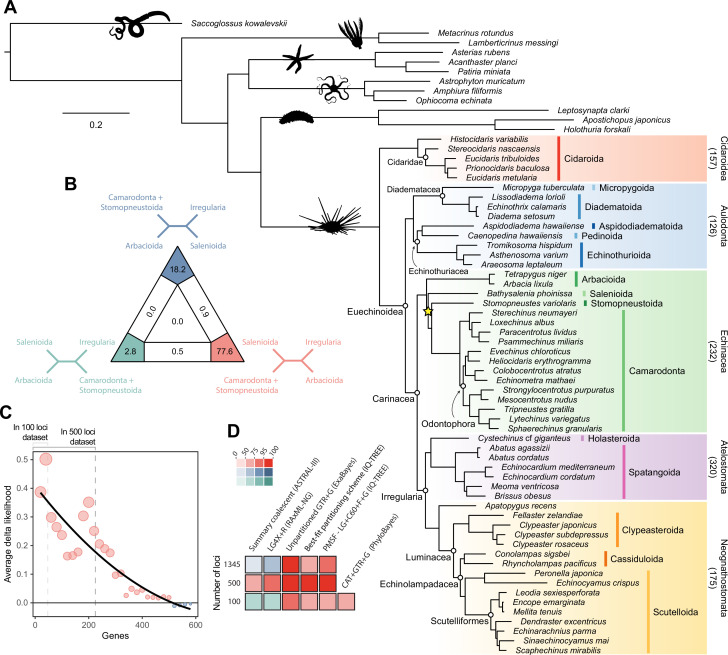
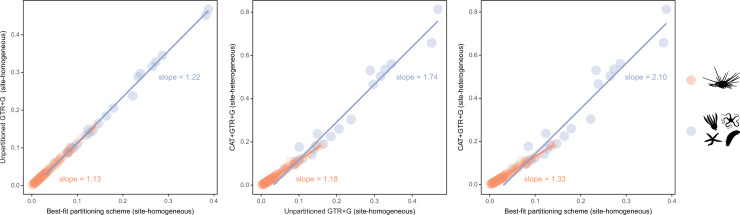
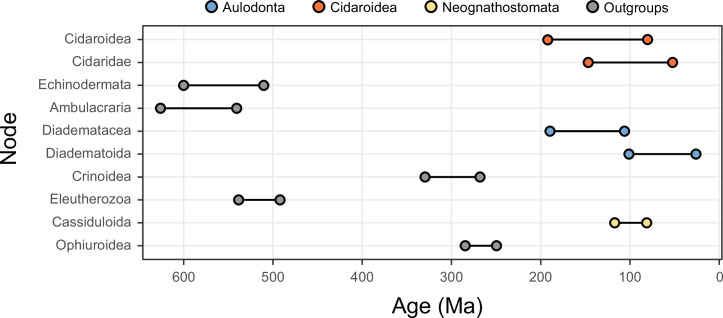
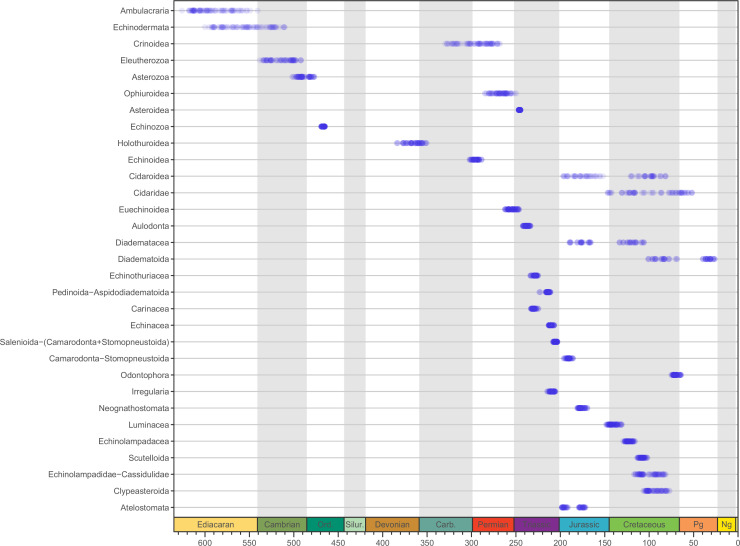
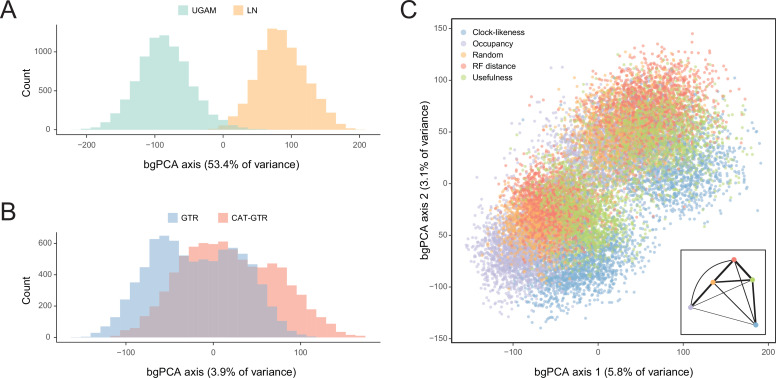
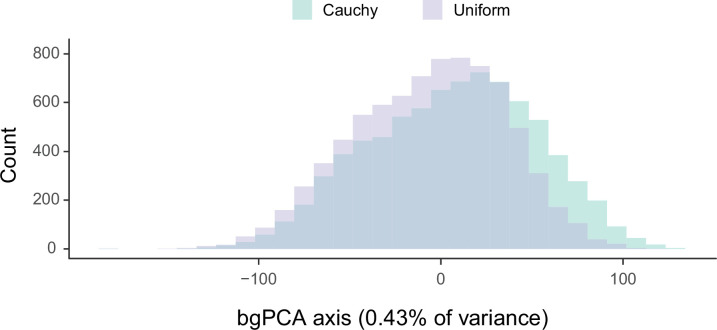
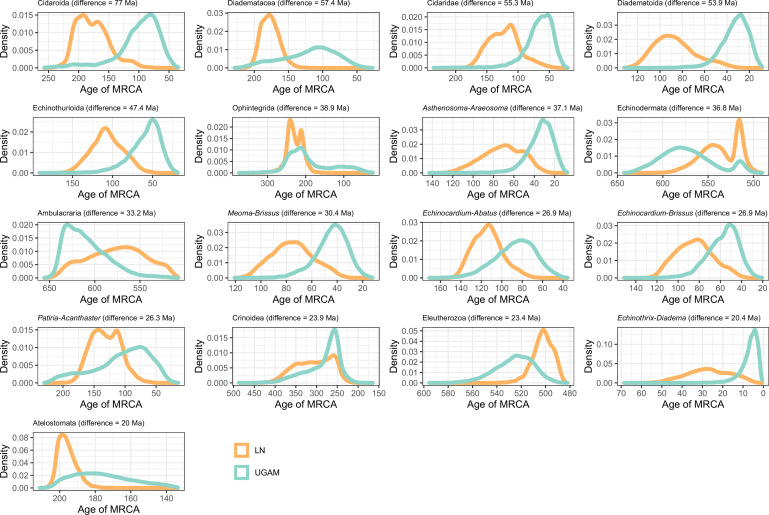
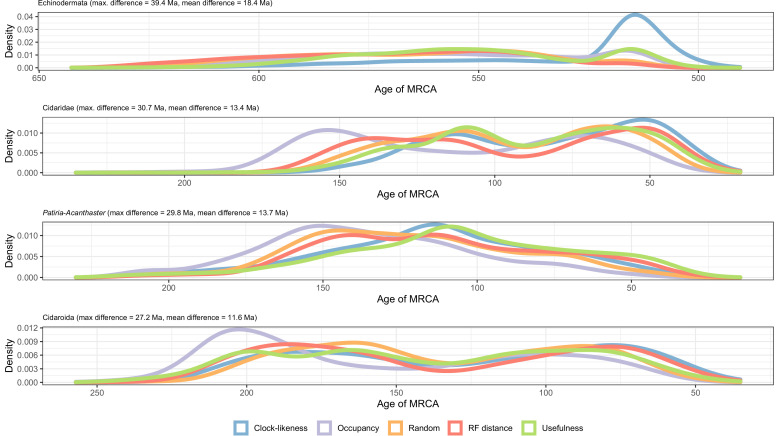
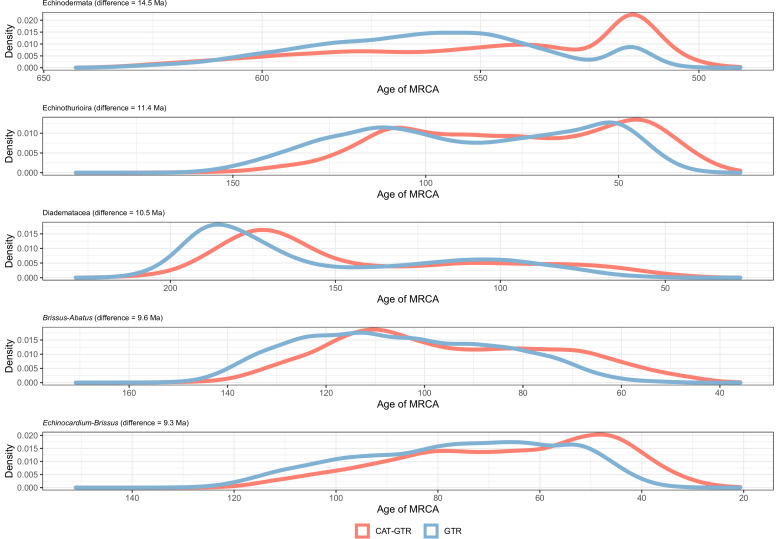
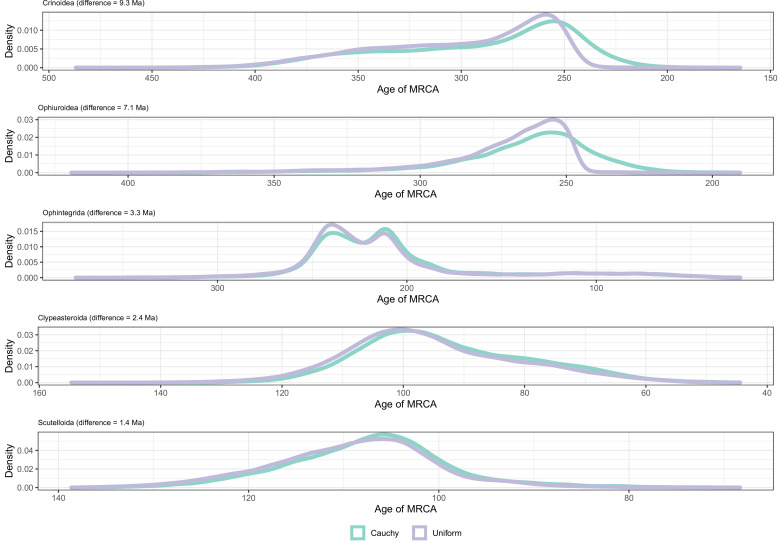
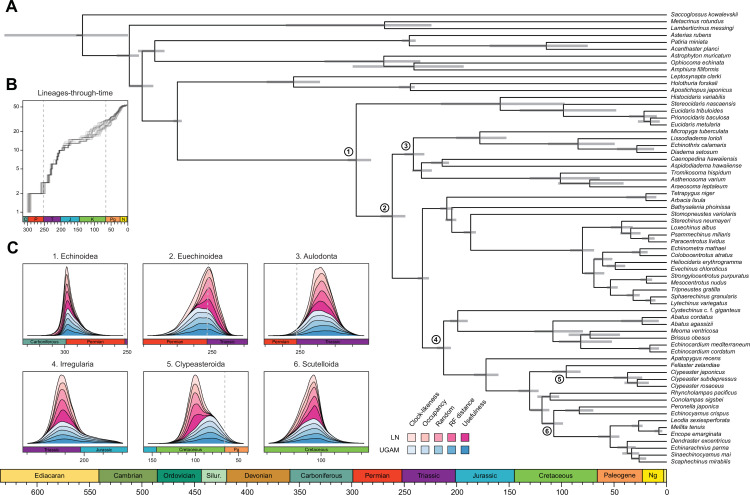
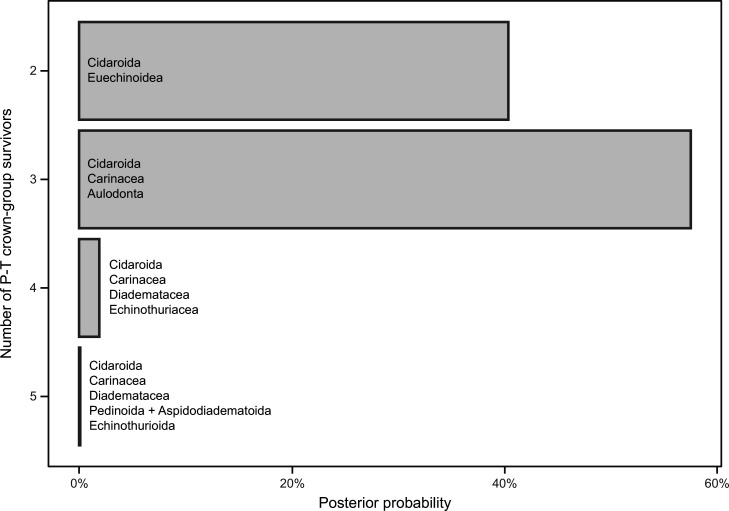
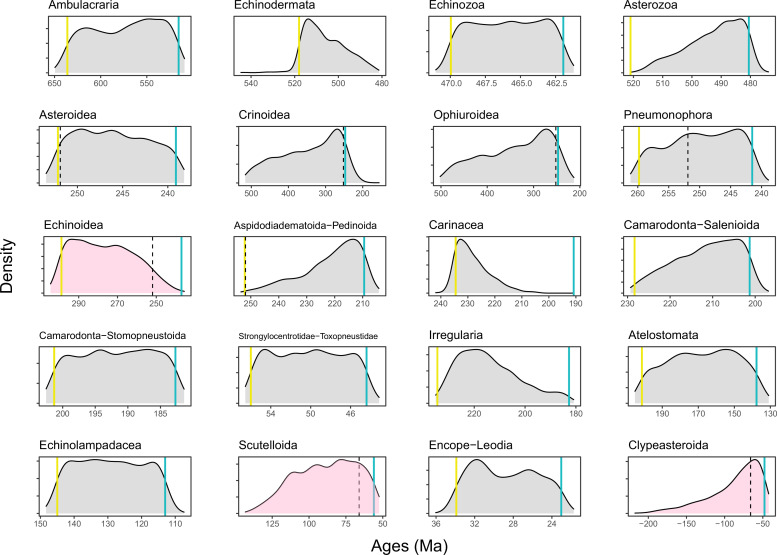
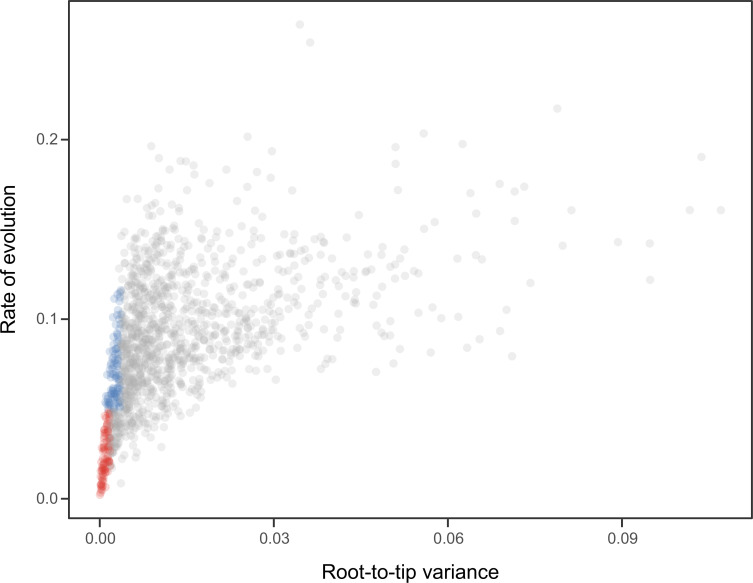
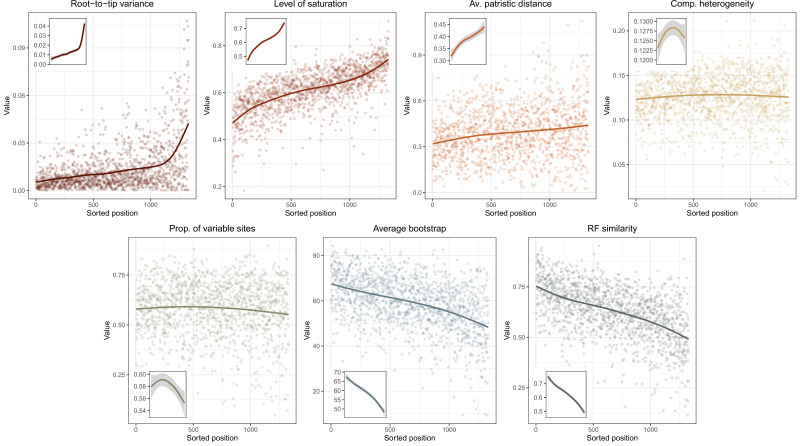
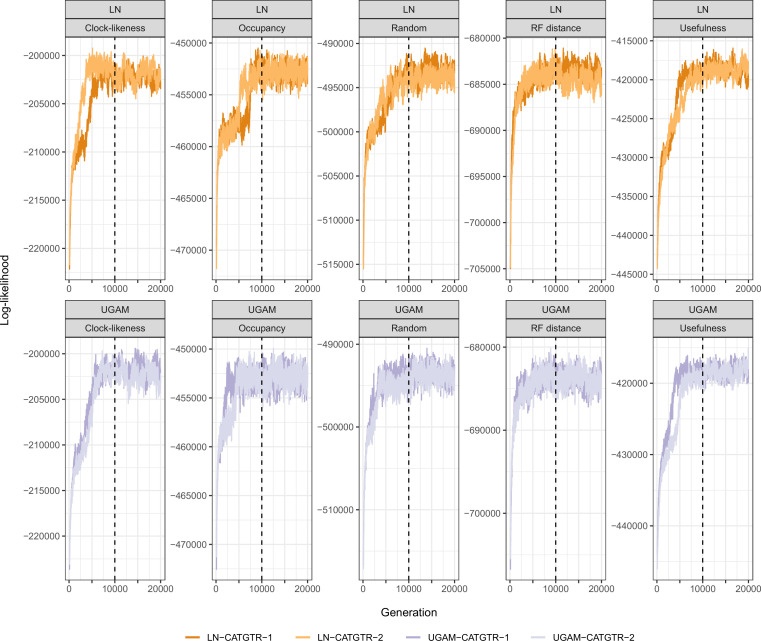
Echinoids are key components of modern marine ecosystems. Despite a remarkable fossil record, the emergence of their crown group is documented by few specimens of unclear affinities, rendering their early history uncertain. The origin of sand dollars, one of its most distinctive clades, is also unclear due to an unstable phylogenetic context. We employ 18 novel genomes and transcriptomes to build a phylogenomic dataset with a near-complete sampling of major lineages. With it, we revise the phylogeny and divergence times of echinoids, and place their history within the broader context of echinoderm evolution. We also introduce the concept of a chronospace - a multidimensional representation of node ages - and use it to explore methodological decisions involved in time calibrating phylogenies. We find the choice of clock model to have the strongest impact on divergence times, while the use of site-heterogeneous models and alternative node prior distributions show minimal effects. The choice of loci has an intermediate impact, affecting mostly deep Paleozoic nodes, for which clock-like genes recover dates more congruent with fossil evidence. Our results reveal that crown group echinoids originated in the Permian and diversified rapidly in the Triassic, despite the relative lack of fossil evidence for this early diversification. We also clarify the relationships between sand dollars and their close relatives and confidently date their origins to the Cretaceous, implying ghost ranges spanning approximately 50 million years, a remarkable discrepancy with their rich fossil record.
Keywords: divergence time estimation; echinoidea; evolutionary biology; phylogenomics; sand dollars; sea urchins; site-heterogeneous models; time calibration.
© 2022, Mongiardino Koch et al.
Conflict of interest statement
NM, JT, AH, MM, AA, SC, FA, OB, AK, RM, GR No competing interests declared
Figures

References
-
- Ali MSM. The paleogeographic distribution of Clypeaster (Echinoidea) during the Cenozoic Era. Neues Jahrb. Für Geol. Und Paläontologie Monatshefte. 1983;8:449–464.
-
- Barras CG. Morphological innovation associated with the expansion of atelostomate irregular echinoids into fine-grained sediments during the Jurassic. Palaeogeography, Palaeoclimatology, Palaeoecology. 2008;263:44–57. doi: 10.1016/j.palaeo.2008.01.026. - DOI
-
- Benton M, Donoghue P, Vinther J, Asher R, Friedman M, Near T. Constraints on the timescale of animal evolutionary history. Palaeontologia Electronica. 2015;18:1–106. doi: 10.26879/424. - DOI
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
