

DNA repair defects in cancer and therapeutic opportunities
- PMID: 35318271
- PMCID: PMC8973847
- DOI: 10.1101/gad.349431.122
DNA repair defects in cancer and therapeutic opportunities
Abstract
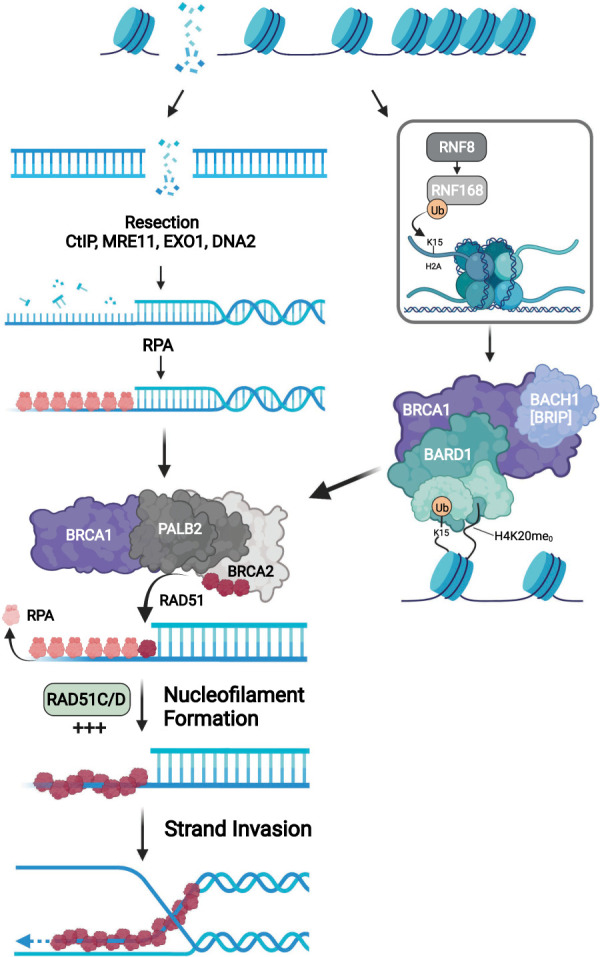
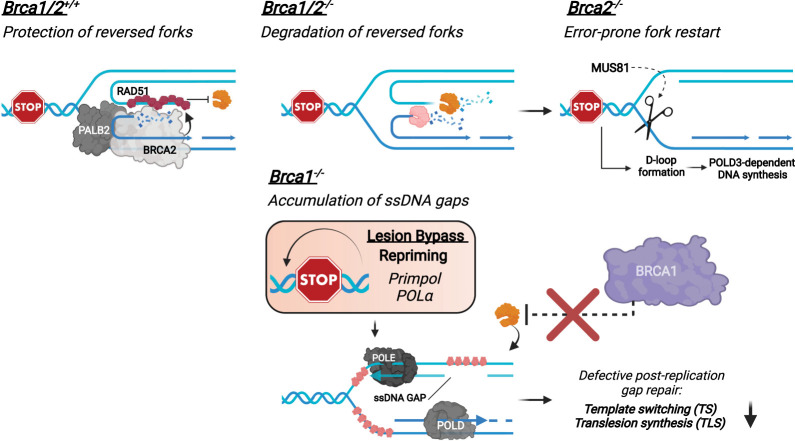
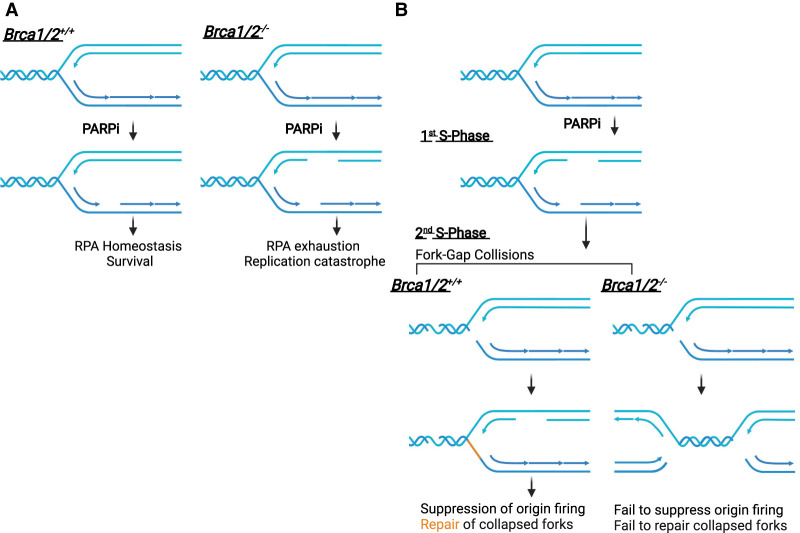
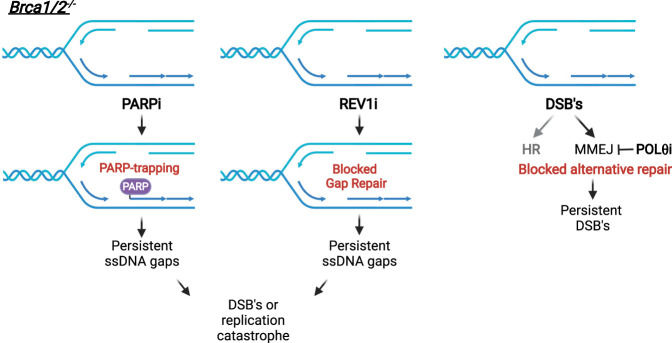
DNA repair and DNA damage signaling pathways are critical for the maintenance of genomic stability. Defects of DNA repair and damage signaling contribute to tumorigenesis, but also render cancer cells vulnerable to DNA damage and reliant on remaining repair and signaling activities. Here, we review the major classes of DNA repair and damage signaling defects in cancer, the genomic instability that they give rise to, and therapeutic strategies to exploit the resulting vulnerabilities. Furthermore, we discuss the impacts of DNA repair defects on both targeted therapy and immunotherapy, and highlight emerging principles for targeting DNA repair defects in cancer therapy.
Keywords: ATM; ATR; BRCA; DNA damage; DNA repair; PARP; cancer; genomic instability; immunotherapy; targeted therapy.
© 2022 Hopkins et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Abbott JM, Zhou Q, Esquer H, Pike L, Broneske TP, Rinaldetti S, Abraham AD, Ramirez DA, Lunghofer PJ, Pitts TM, et al. 2020. First-in-class inhibitors of oncogenic CHD1L with preclinical activity against colorectal cancer. Mol Cancer Ther 19: 1598–1612. 10.1158/1535-7163.MCT-20-0106 - DOI - PMC - PubMed
-
- Adam S, Rossi SE, Moatti N, De Marco Zompit M, Xue Y, Ng TF, Álvarez-Quilón A, Desjardins J, Bhaskaran V, Martino G, et al. 2021. The CIP2A–TOPBP1 axis safeguards chromosome stability and is a synthetic lethal target for BRCA-mutated cancer. Nat Cancer 2: 1357–1371. 10.1038/s43018-021-00266-w - DOI - PubMed
-
- Álvarez-Quilón A, Wojtaszek JL, Mathieu MC, Patel T, Appel CD, Hustedt N, Rossi SE, Wallace BD, Setiaputra D, Adam S, et al. 2020. Endogenous DNA 3′ blocks Are vulnerabilities for BRCA1 and BRCA2 deficiency and are reversed by the APE2 nuclease. Mol Cell 78: 1152–1165.e8. 10.1016/j.molcel.2020.05.021 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous