

SARS-CoV-2, SARS-CoV, and MERS-CoV encode circular RNAs of spliceosome-independent origin
- PMID: 35318674
- PMCID: PMC9088628
- DOI: 10.1002/jmv.27734
SARS-CoV-2, SARS-CoV, and MERS-CoV encode circular RNAs of spliceosome-independent origin
Abstract
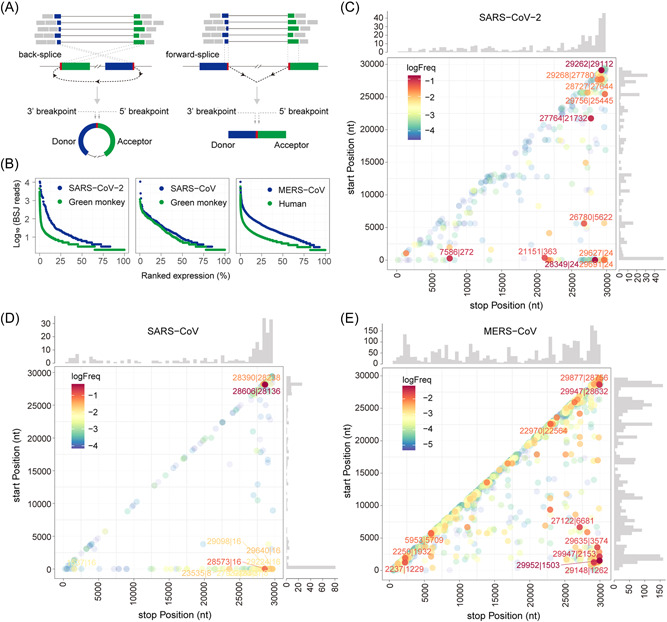
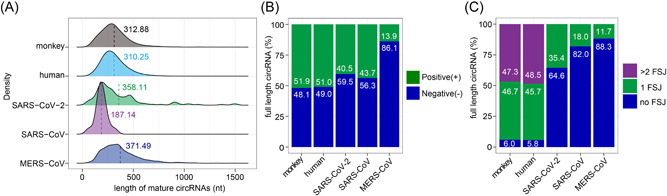
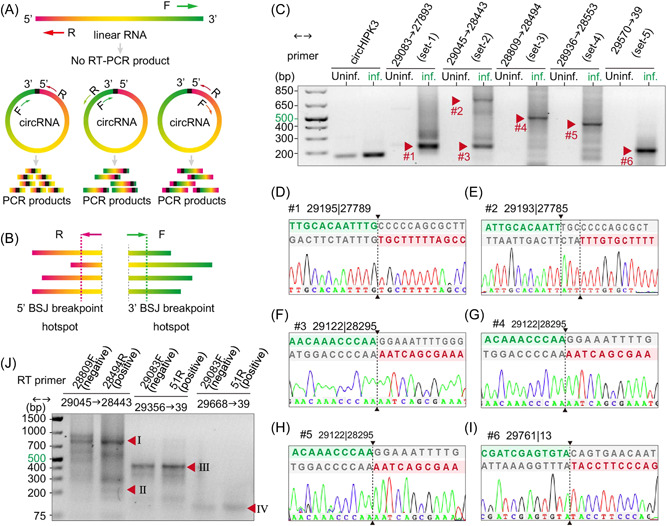
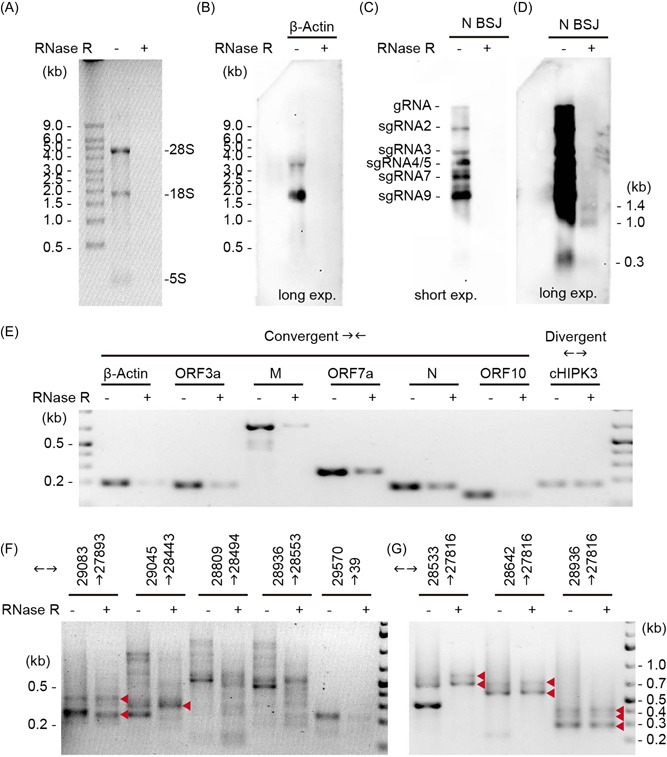
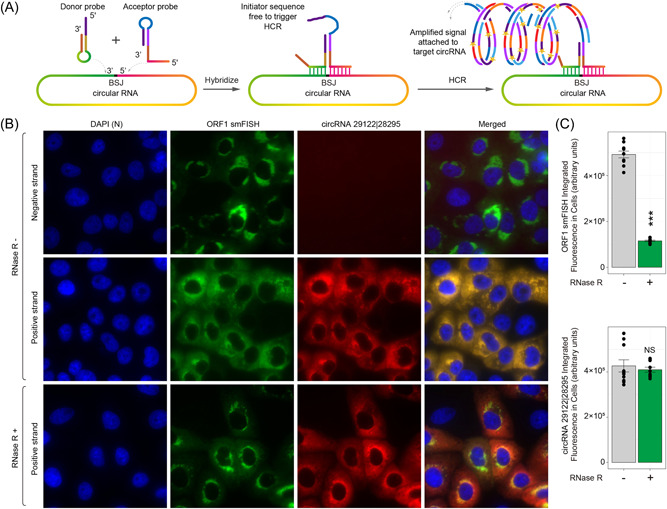
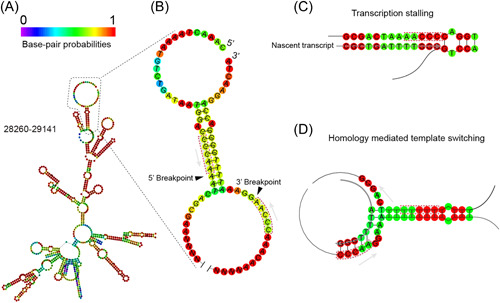
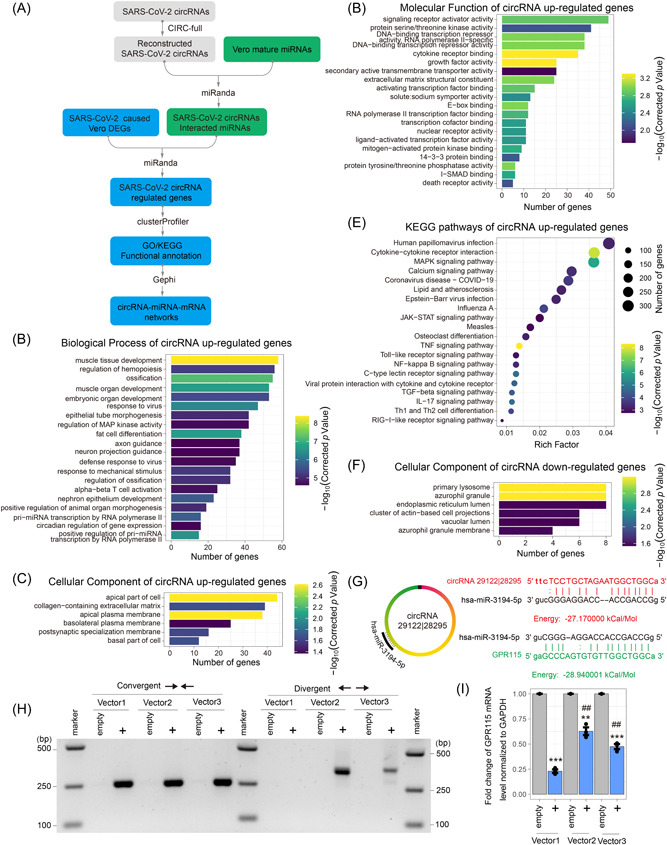
Circular RNAs (circRNAs) are a newly recognized component of the transcriptome with critical roles in autoimmune diseases and viral pathogenesis. To address the importance of circRNA in RNA viral transcriptome, we systematically identified and characterized circRNAs encoded by the RNA genomes of betacoronaviruses using both bioinformatical and experimental approaches. We predicted 351, 224, and 2764 circRNAs derived from severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), SARS-CoV, and Middle East respiratory syndrome coronavirus, respectively. We experimentally identified 75 potential SARS-CoV-2 circRNAs from RNA samples extracted from SARS-CoV-2-infected Vero E6 cells. A systematic comparison of viral and host circRNA features, including abundance, strand preference, length distribution, circular exon numbers, and breakpoint sequences, demonstrated that coronavirus-derived circRNAs had a spliceosome-independent origin. We further showed that back-splice junctions (BSJs) captured by inverse reverse-transcription polymerase chain reaction have different level of resistance to RNase R. Through northern blotting with a BSJ-spanning probe targeting N gene, we identified three RNase R-resistant bands that represent SARS-CoV-2 circRNAs that are detected cytoplasmic by single-molecule and amplified fluorescence in situ hybridization assays. Lastly, analyses of 169 sequenced BSJs showed that both back-splice and forward-splice junctions were flanked by homologous and reverse complementary sequences, including but not limited to the canonical transcriptional regulatory sequences. Our findings highlight circRNAs as an important component of the coronavirus transcriptome, offer important evaluation of bioinformatic tools in the analysis of circRNAs from an RNA genome, and shed light on the mechanism of discontinuous RNA synthesis.
Keywords: RNA biology; SARS-CoV-2; circular RNA; coronavirus; virology.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
The authors declare noconflicts of interest.
Figures
Update of
-
Circular RNA profiling reveals abundant and diverse circRNAs of SARS-CoV-2, SARS-CoV and MERS-CoV origin.bioRxiv [Preprint]. 2020 Dec 8:2020.12.07.415422. doi: 10.1101/2020.12.07.415422. bioRxiv. 2020. Update in: J Med Virol. 2022 Jul;94(7):3203-3222. doi: 10.1002/jmv.27734. PMID: 33330860 Free PMC article. Updated. Preprint.
Similar articles
-
Circular RNA profiling reveals abundant and diverse circRNAs of SARS-CoV-2, SARS-CoV and MERS-CoV origin.bioRxiv [Preprint]. 2020 Dec 8:2020.12.07.415422. doi: 10.1101/2020.12.07.415422. bioRxiv. 2020. Update in: J Med Virol. 2022 Jul;94(7):3203-3222. doi: 10.1002/jmv.27734. PMID: 33330860 Free PMC article. Updated. Preprint.
-
Circular RNAs Represent a Novel Class of Human Cytomegalovirus Transcripts.Microbiol Spectr. 2022 Jun 29;10(3):e0110622. doi: 10.1128/spectrum.01106-22. Epub 2022 May 23. Microbiol Spectr. 2022. PMID: 35604147 Free PMC article.
-
hnRNP C modulates MERS-CoV and SARS-CoV-2 replication by governing the expression of a subset of circRNAs and cognitive mRNAs.Emerg Microbes Infect. 2022 Dec;11(1):519-531. doi: 10.1080/22221751.2022.2032372. Emerg Microbes Infect. 2022. PMID: 35060842 Free PMC article.
-
Review on circular RNAs and new insights into their roles in cancer.Comput Struct Biotechnol J. 2021 Jan 22;19:910-928. doi: 10.1016/j.csbj.2021.01.018. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 33598105 Free PMC article. Review.
-
Circular RNAs as emerging regulators in COVID-19 pathogenesis and progression.Front Immunol. 2022 Nov 9;13:980231. doi: 10.3389/fimmu.2022.980231. eCollection 2022. Front Immunol. 2022. PMID: 36439162 Free PMC article. Review.
Cited by
-
Virus-derived circular RNAs populate hepatitis C virus-infected cells.Proc Natl Acad Sci U S A. 2024 Feb 13;121(7):e2313002121. doi: 10.1073/pnas.2313002121. Epub 2024 Feb 6. Proc Natl Acad Sci U S A. 2024. PMID: 38319965 Free PMC article.
-
Virus-Encoded Circular RNAs: Role and Significance in Viral Infections.Int J Mol Sci. 2023 Nov 20;24(22):16547. doi: 10.3390/ijms242216547. Int J Mol Sci. 2023. PMID: 38003737 Free PMC article. Review.
-
The Internal Ribosome Entry Site in Circular RNAs.Adv Exp Med Biol. 2025;1485:131-148. doi: 10.1007/978-981-96-9428-0_9. Adv Exp Med Biol. 2025. PMID: 40886274 Review.
-
HMCDA: a novel method based on the heterogeneous graph neural network and metapath for circRNA-disease associations prediction.BMC Bioinformatics. 2023 Sep 11;24(1):335. doi: 10.1186/s12859-023-05441-7. BMC Bioinformatics. 2023. PMID: 37697297 Free PMC article.
-
Identification and characterization of Varicella Zoster Virus circular RNA in lytic infection.Nat Commun. 2024 Jun 10;15(1):4932. doi: 10.1038/s41467-024-49112-4. Nat Commun. 2024. PMID: 38858365 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
