

AKR1D1 knockout mice develop a sex-dependent metabolic phenotype
- PMID: 35318963
- PMCID: PMC9086936
- DOI: 10.1530/JOE-21-0280
AKR1D1 knockout mice develop a sex-dependent metabolic phenotype
Abstract
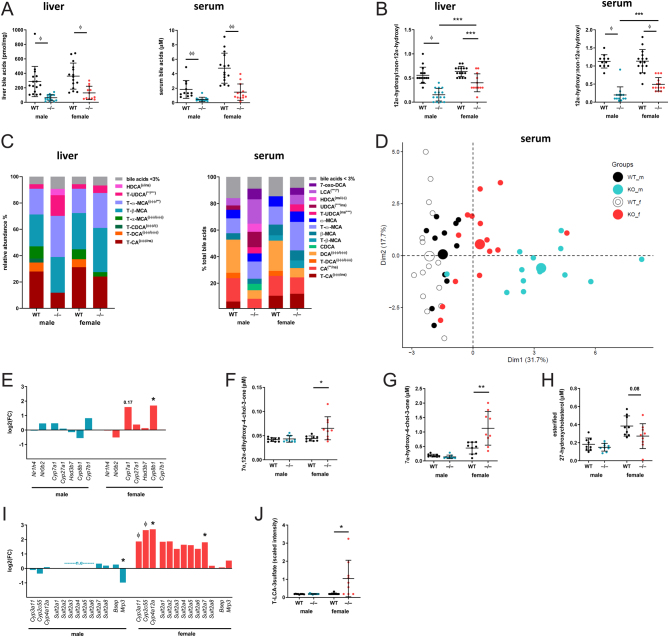
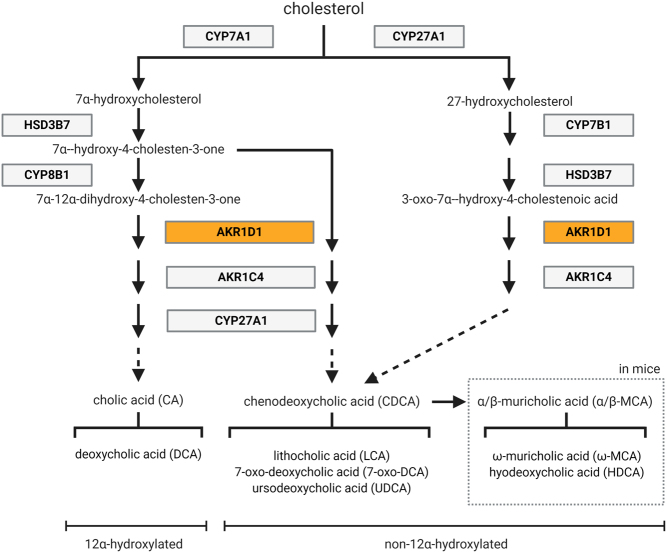
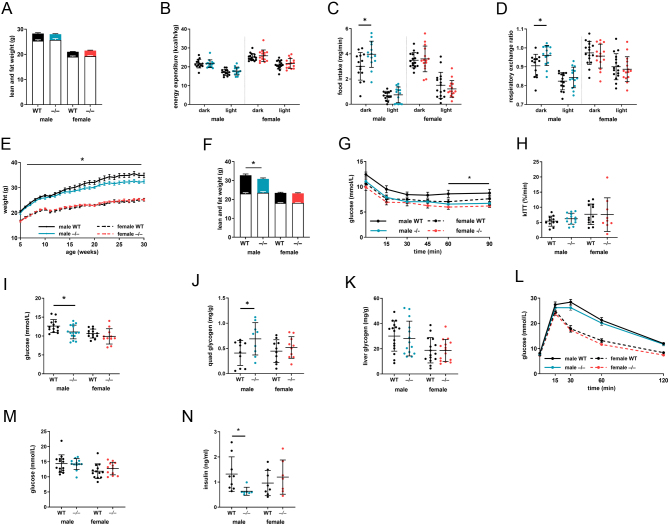
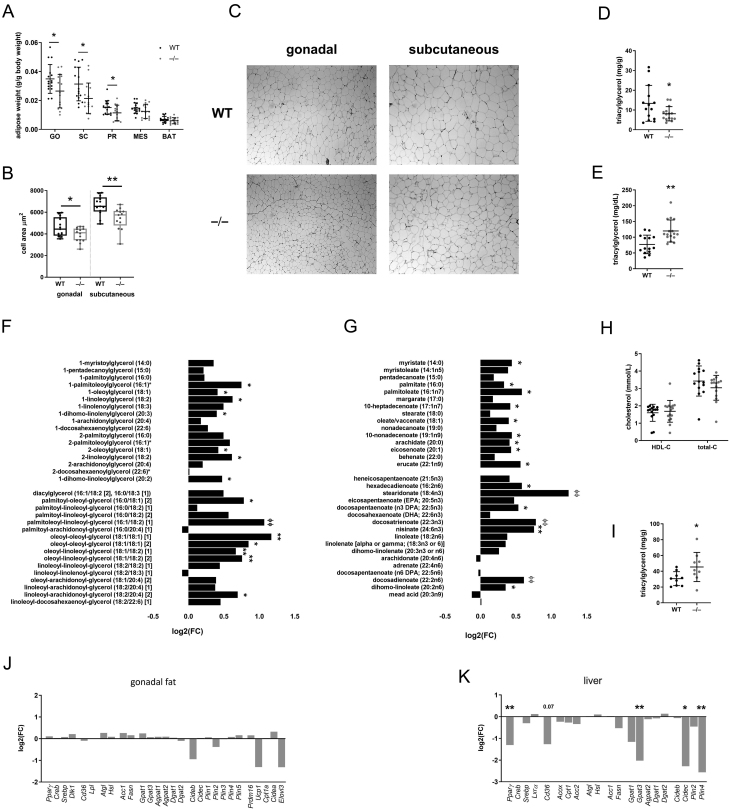
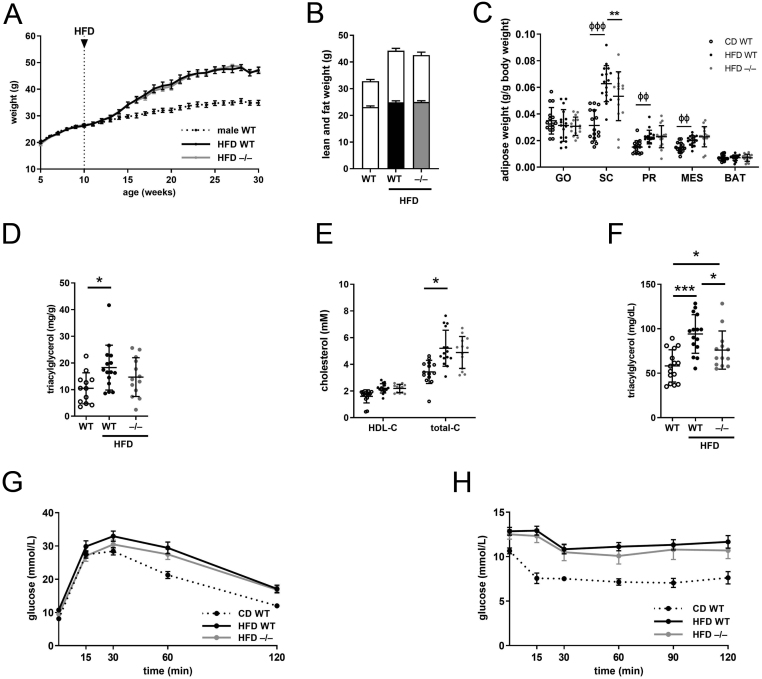
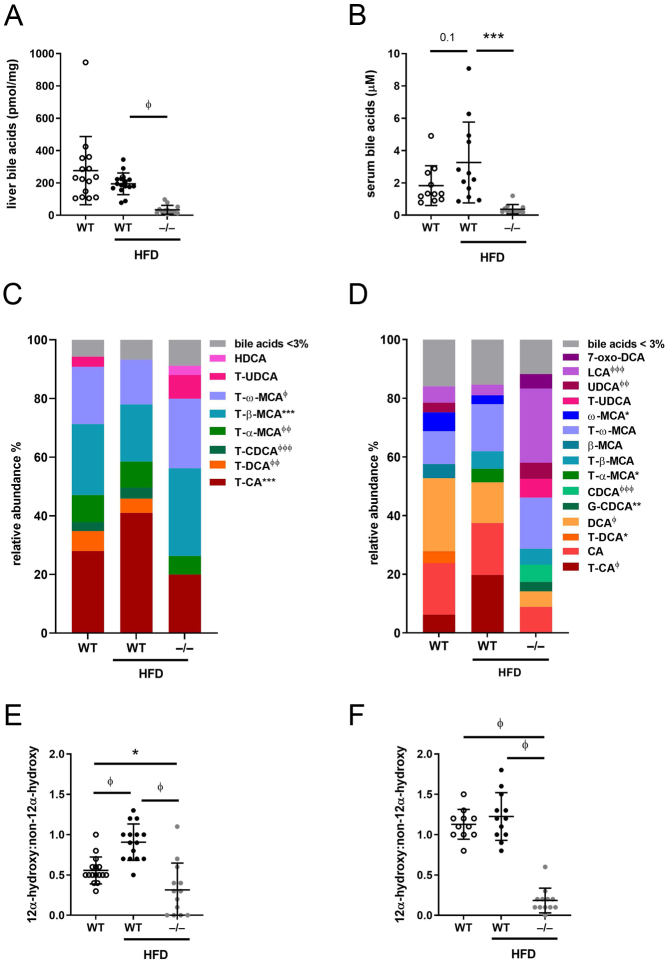
Steroid 5β-reductase (AKR1D1) plays important role in hepatic bile acid synthesis and glucocorticoid clearance. Bile acids and glucocorticoids are potent metabolic regulators, but whether AKR1D1 controls metabolic phenotype in vivo is unknown. Akr1d1-/- mice were generated on a C57BL/6 background. Liquid chromatography/mass spectrometry, metabolomic and transcriptomic approaches were used to determine effects on glucocorticoid and bile acid homeostasis. Metabolic phenotypes including body weight and composition, lipid homeostasis, glucose tolerance and insulin tolerance were evaluated. Molecular changes were assessed by RNA-Seq and Western blotting. Male Akr1d1-/- mice were challenged with a high fat diet (60% kcal from fat) for 20 weeks. Akr1d1-/- mice had a sex-specific metabolic phenotype. At 30 weeks of age, male, but not female, Akr1d1-/- mice were more insulin tolerant and had reduced lipid accumulation in the liver and adipose tissue yet had hypertriglyceridemia and increased intramuscular triacylglycerol. This phenotype was associated with sexually dimorphic changes in bile acid metabolism and composition but without overt effects on circulating glucocorticoid levels or glucocorticoid-regulated gene expression in the liver. Male Akr1d1-/- mice were not protected against diet-induced obesity and insulin resistance. In conclusion, this study shows that AKR1D1 controls bile acid homeostasis in vivo and that altering its activity can affect insulin tolerance and lipid homeostasis in a sex-dependent manner.
Keywords: bile acid; chenodeoxycholic acid; cholic acid; cortisol; metabolic syndrome; non-alcoholic fatty liver disease; steroid.
Figures
References
-
- Abdelkarim M, Caron S, Duhem C, Prawitt J, Dumont J, Lucas A, Bouchaert E, Briand O, Brozek J, Kuipers Fet al.2010The farnesoid X receptor regulates adipocyte differentiation and function by promoting peroxisome proliferator-activated receptor-gamma and interfering with the Wnt/beta-catenin pathways. Journal of Biological Chemistry 28536759–36767. (10.1074/jbc.M110.166231) - DOI - PMC - PubMed
-
- Bertaggia E, Jensen KK, Castro-Perez J, Xu Y, Di Paolo G, Chan RB, Wang L, Haeusler RA.2017 Cyp8b1 ablation prevents Western diet-induced weight gain and hepatic steatosis because of impaired fat absorption. American Journal of Physiology: Endocrinology and Metabolism 313E121–E133. (10.1152/ajpendo.00409.2016) - DOI - PMC - PubMed
-
- Cariou B, Van Harmelen K, Duran-Sandoval D, Van Dijk TH, Grefhorst A, Abdelkarim M, Caron S, Torpier G, Fruchart JC, Gonzalez FJet al.2006The farnesoid X receptor modulates adiposity and peripheral insulin sensitivity in mice. Journal of Biological Chemistry 28111039–11049. (10.1074/jbc.M510258200) - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
