

Temporal control of the integrated stress response by a stochastic molecular switch
- PMID: 35319985
- PMCID: PMC8942376
- DOI: 10.1126/sciadv.abk2022
Temporal control of the integrated stress response by a stochastic molecular switch
Abstract
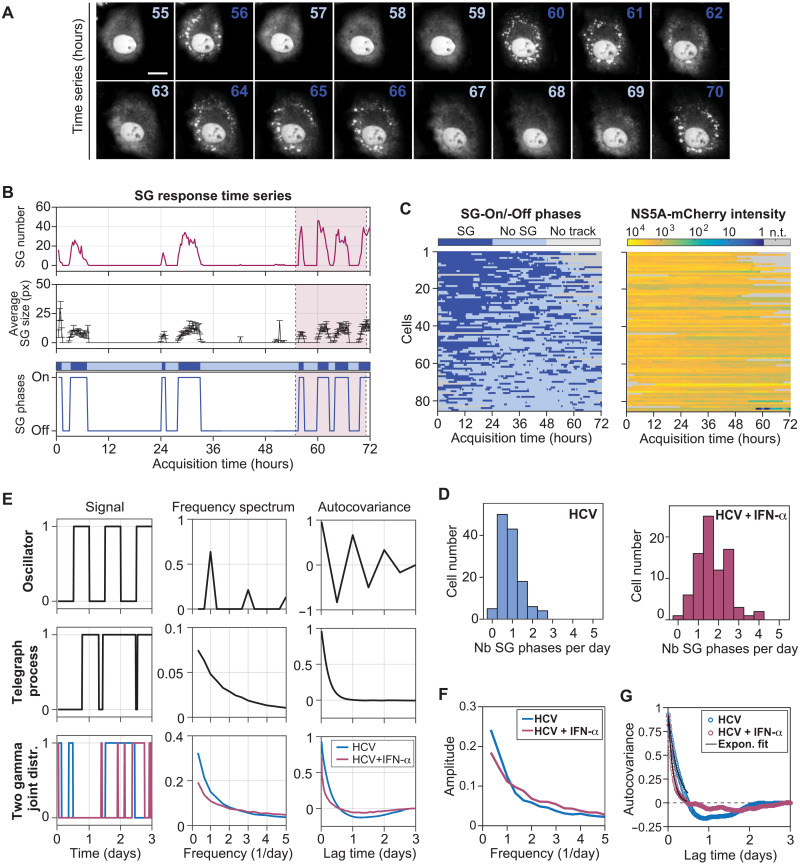
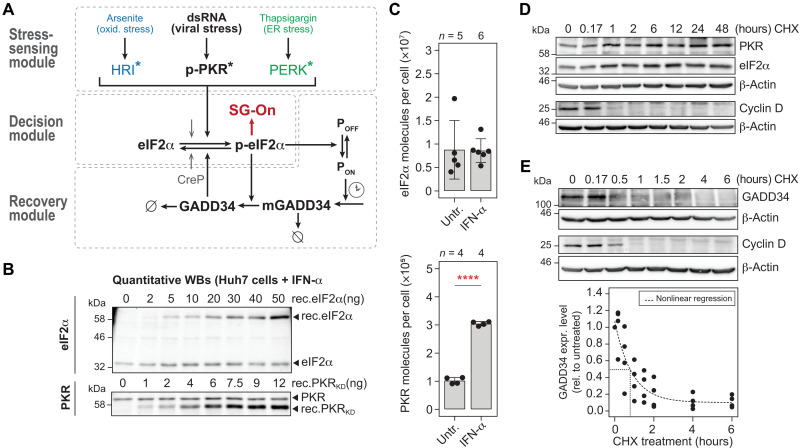
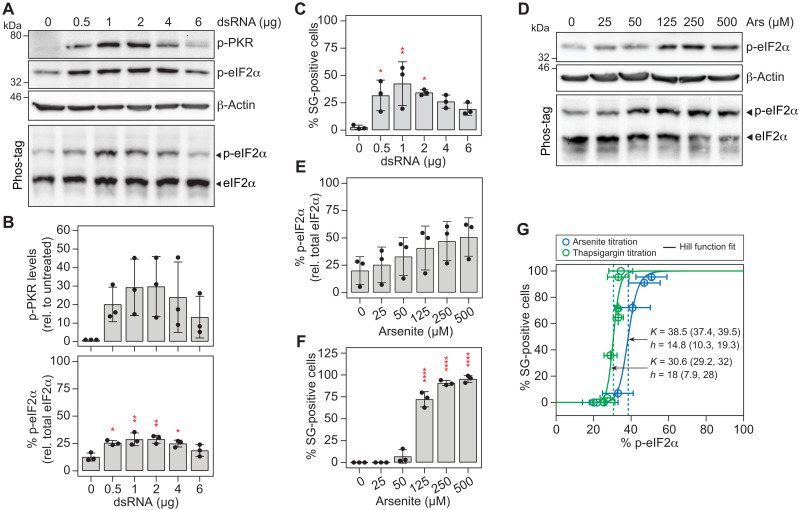
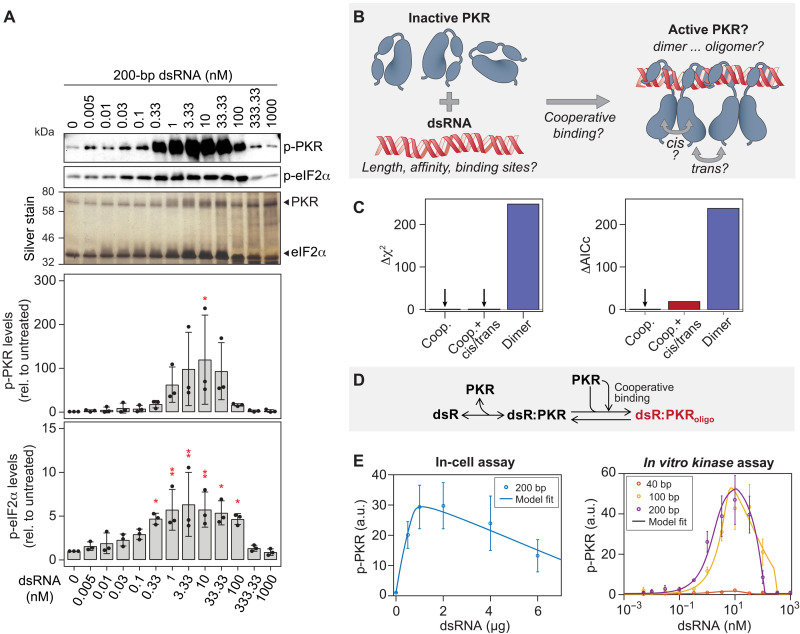
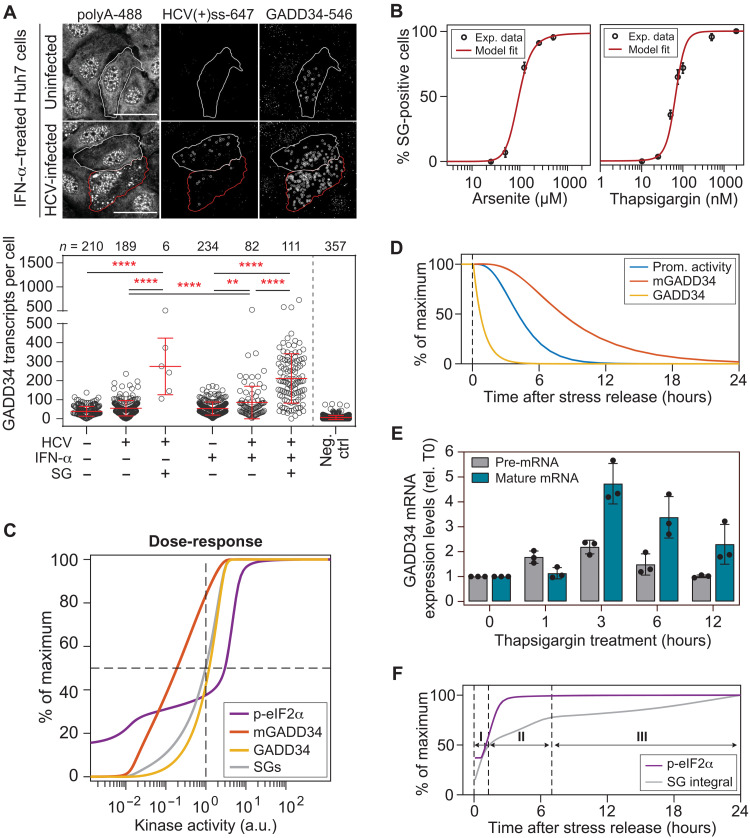
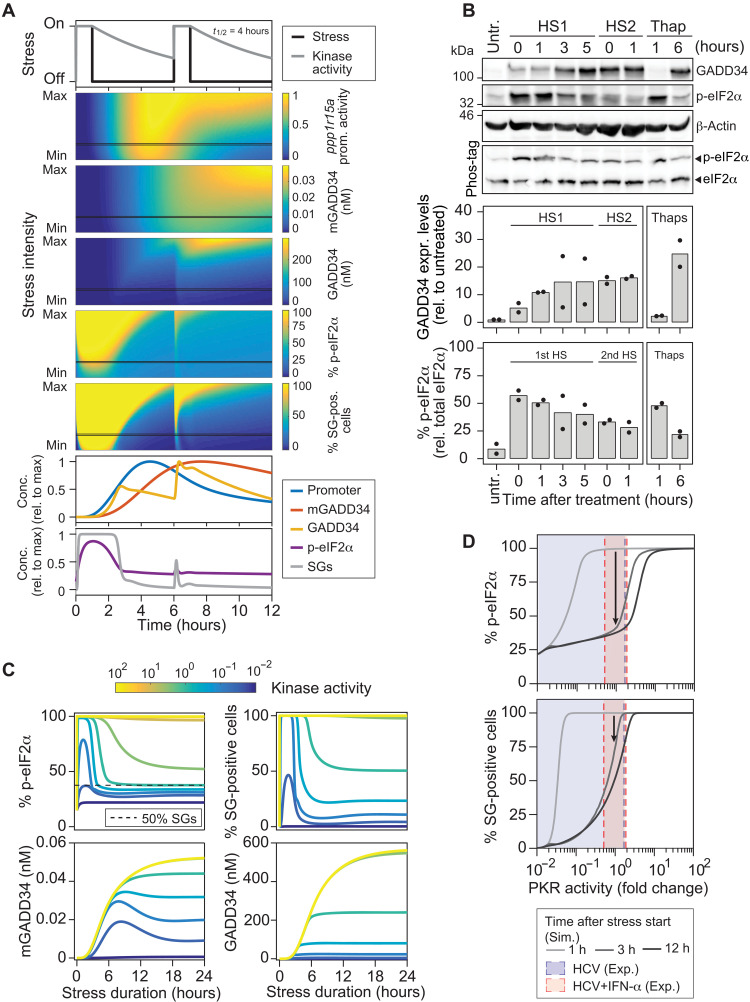
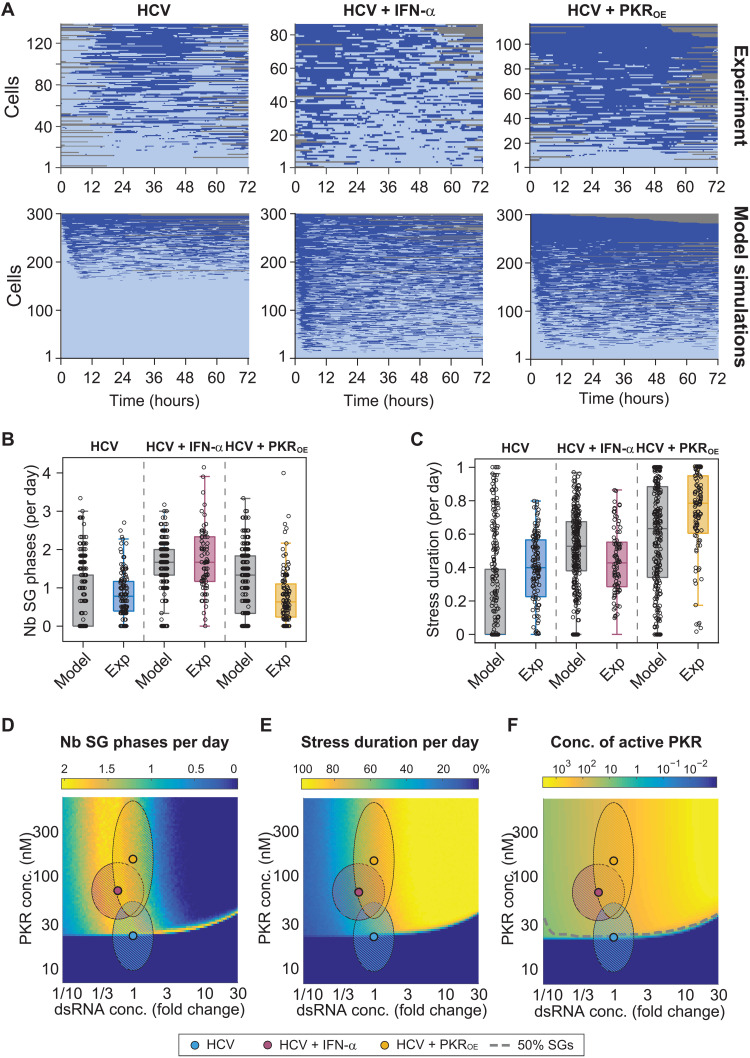
Stress granules (SGs) are formed in the cytosol as an acute response to environmental cues and activation of the integrated stress response (ISR), a central signaling pathway controlling protein synthesis. Using chronic virus infection as stress model, we previously uncovered a unique temporal control of the ISR resulting in recurrent phases of SG assembly and disassembly. Here, we elucidate the molecular network generating this fluctuating stress response by integrating quantitative experiments with mathematical modeling and find that the ISR operates as a stochastic switch. Key elements controlling this switch are the cooperative activation of the stress-sensing kinase PKR, the ultrasensitive response of SG formation to the phosphorylation of the translation initiation factor eIF2α, and negative feedback via GADD34, a stress-induced subunit of protein phosphatase 1. We identify GADD34 messenger RNA levels as the molecular memory of the ISR that plays a central role in cell adaptation to acute and chronic stress.
Figures
Similar articles
-
Mouse Norovirus Infection Arrests Host Cell Translation Uncoupled from the Stress Granule-PKR-eIF2α Axis.mBio. 2019 Jun 18;10(3):e00960-19. doi: 10.1128/mBio.00960-19. mBio. 2019. PMID: 31213553 Free PMC article.
-
Defining the Role of Stress Granules in Innate Immune Suppression by the Herpes Simplex Virus 1 Endoribonuclease VHS.J Virol. 2018 Jul 17;92(15):e00829-18. doi: 10.1128/JVI.00829-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29793959 Free PMC article.
-
Hiding in Plain Sight: Formation and Function of Stress Granules During Microbial Infection of Mammalian Cells.Front Mol Biosci. 2021 Apr 29;8:647884. doi: 10.3389/fmolb.2021.647884. eCollection 2021. Front Mol Biosci. 2021. PMID: 33996904 Free PMC article. Review.
-
The PERK/PKR-eIF2α Pathway Negatively Regulates Porcine Hemagglutinating Encephalomyelitis Virus Replication by Attenuating Global Protein Translation and Facilitating Stress Granule Formation.J Virol. 2022 Jan 12;96(1):e0169521. doi: 10.1128/JVI.01695-21. Epub 2021 Oct 13. J Virol. 2022. PMID: 34643429 Free PMC article.
-
The Integrated Stress Response and Phosphorylated Eukaryotic Initiation Factor 2α in Neurodegeneration.J Neuropathol Exp Neurol. 2020 Feb 1;79(2):123-143. doi: 10.1093/jnen/nlz129. J Neuropathol Exp Neurol. 2020. PMID: 31913484 Free PMC article. Review.
Cited by
-
Dual Nature of Mitochondrial Integrated Stress Response: Molecular Switches from Protection to Pathology.Genes (Basel). 2025 Aug 13;16(8):957. doi: 10.3390/genes16080957. Genes (Basel). 2025. PMID: 40870005 Free PMC article. Review.
-
Stress granule formation helps to mitigate neurodegeneration.bioRxiv [Preprint]. 2023 Nov 11:2023.11.07.566060. doi: 10.1101/2023.11.07.566060. bioRxiv. 2023. Update in: Nucleic Acids Res. 2024 Sep 9;52(16):9745-9759. doi: 10.1093/nar/gkae655. PMID: 37986813 Free PMC article. Updated. Preprint.
-
SARS-CoV-2 Nucleocapsid Protein Antagonizes GADD34-Mediated Innate Immune Pathway through Atypical Foci.Molecules. 2024 Oct 10;29(20):4792. doi: 10.3390/molecules29204792. Molecules. 2024. PMID: 39459161 Free PMC article.
-
Signaling plasticity in the integrated stress response.Front Cell Dev Biol. 2023 Dec 7;11:1271141. doi: 10.3389/fcell.2023.1271141. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38143923 Free PMC article. Review.
-
Induction of Stress Granules and Developmental Instability of Offspring Phenotype Due to Hypothermia During First Mouse Embryo Cleavage.Int J Mol Sci. 2025 Aug 20;26(16):8060. doi: 10.3390/ijms26168060. Int J Mol Sci. 2025. PMID: 40869380 Free PMC article.
References
-
- Holcik M., Sonenberg N., Translational control in stress and apoptosis. Nat. Rev. Mol. Cell Biol. 6, 318–327 (2005). - PubMed
-
- Meurs E., Chong K., Galabru J., Thomas N. S. B., Kerr I. M., Williams B. R. G., Hovanessian A. G., Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell 62, 379–390 (1990). - PubMed
LinkOut - more resources
Full Text Sources
