

The virota and its transkingdom interactions in the healthy infant gut
- PMID: 35320047
- PMCID: PMC9060457
- DOI: 10.1073/pnas.2114619119
The virota and its transkingdom interactions in the healthy infant gut
Abstract
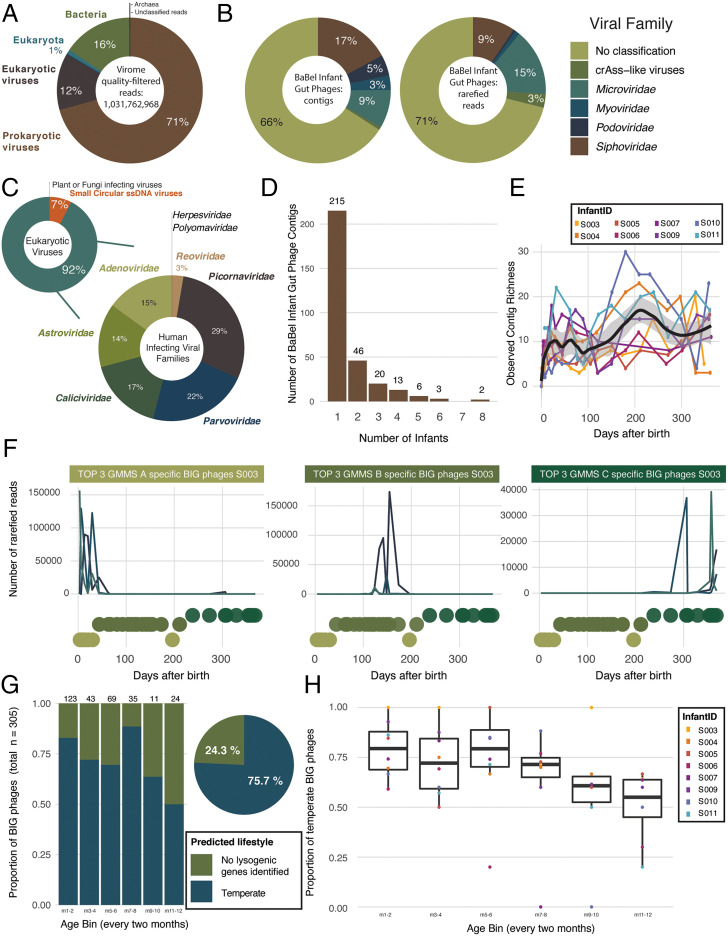
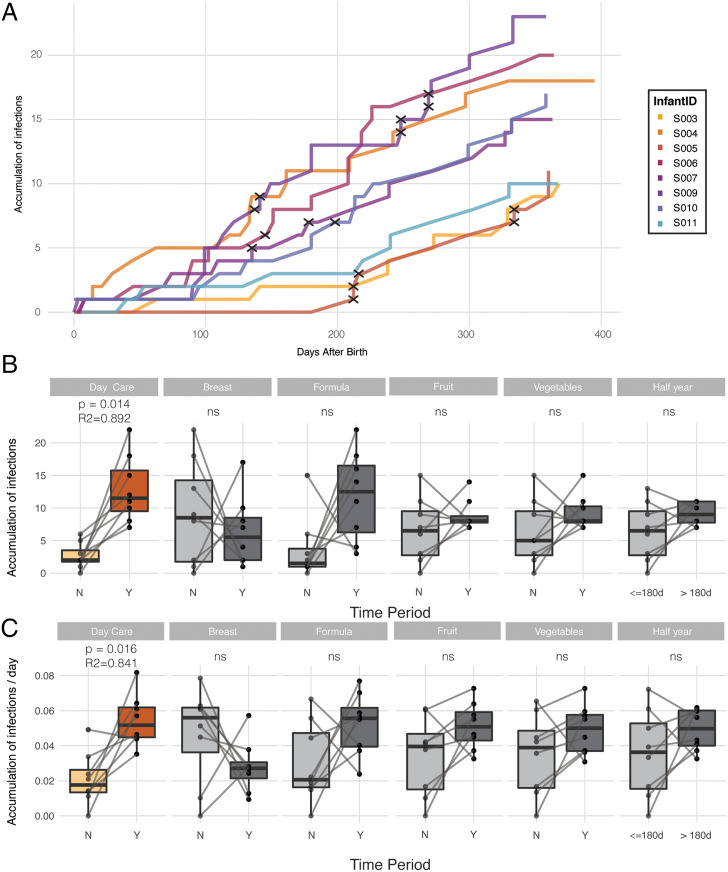
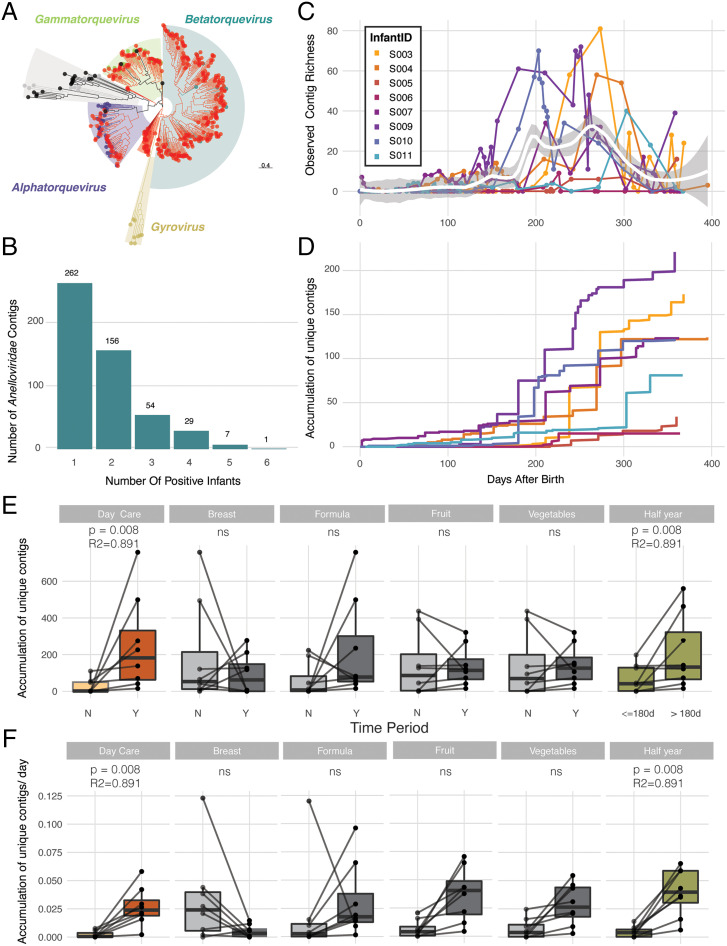
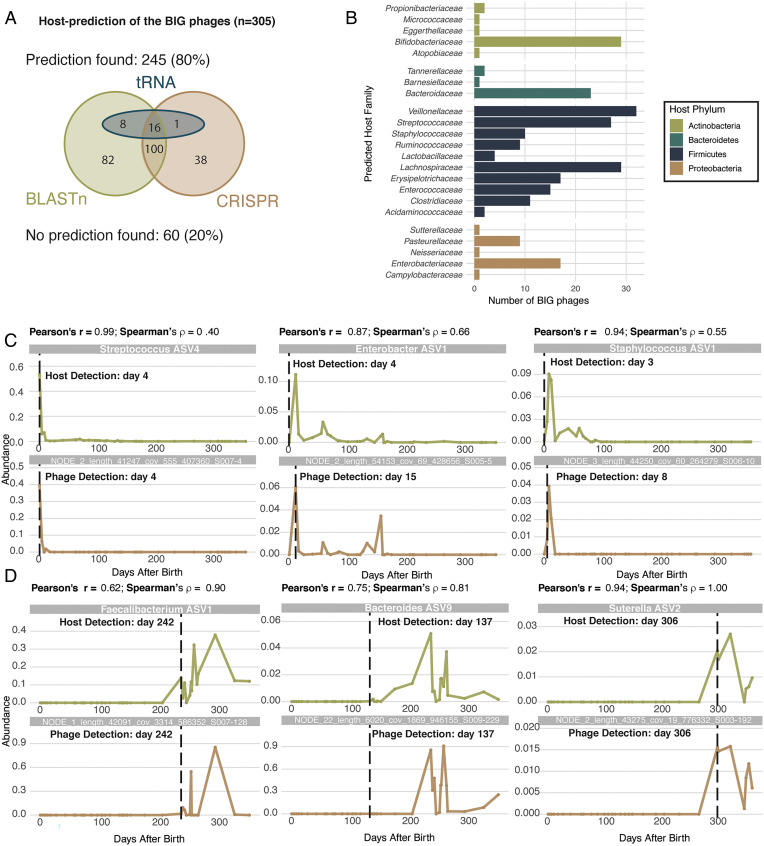
SignificanceMicrobes colonizing the infant gut during the first year(s) of life play an important role in immune system development. We show that after birth the (nearly) sterile gut is rapidly colonized by bacteria and their viruses (phages), which often show a strong cooccurrence. Most viruses infecting the infant do not cause clinical signs and their numbers strongly increase after day-care entrance. The infant diet is clearly reflected by identification of plant-infecting viruses, whereas fungi and parasites are not part of a stable gut microbiota. These temporal high-resolution baseline data about the gut colonization process will be valuable for further investigations of pathogenic viruses, dynamics between phages and their bacterial host, as well as studies investigating infants with a disturbed microbiota.
Keywords: infant; microbiota; transkingdom; virome; virota.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Wang B., Yao M., Lv L., Ling Z., Li L., The human microbiota in health and disease. Engineering 3, 71–82 (2017).
-
- Poyet M., et al. , A library of human gut bacterial isolates paired with longitudinal multiomics data enables mechanistic microbiome research. Nat. Med. 25, 1442–1452 (2019). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
