

MDC1 is essential for G2/M transition and spindle assembly in mouse oocytes
- PMID: 35320416
- PMCID: PMC11071937
- DOI: 10.1007/s00018-022-04241-1
MDC1 is essential for G2/M transition and spindle assembly in mouse oocytes
Abstract
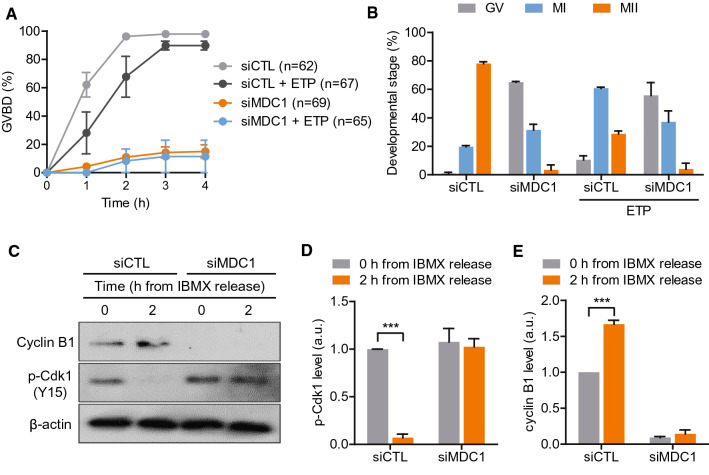
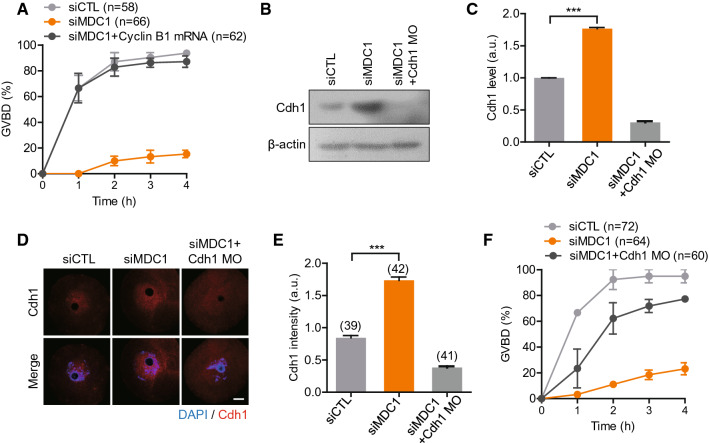
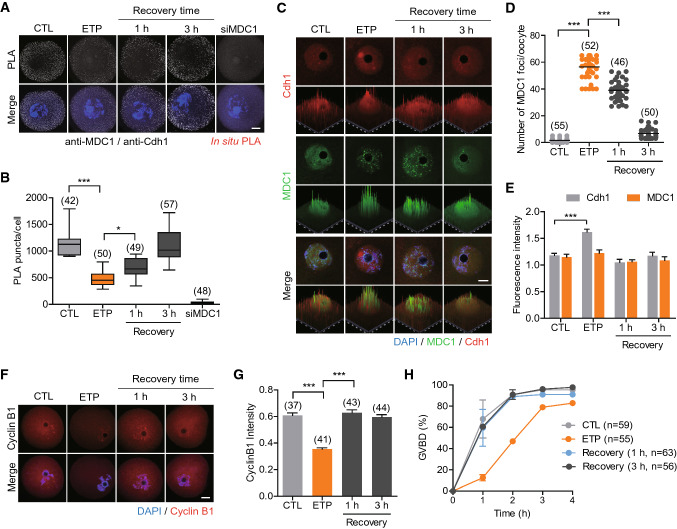
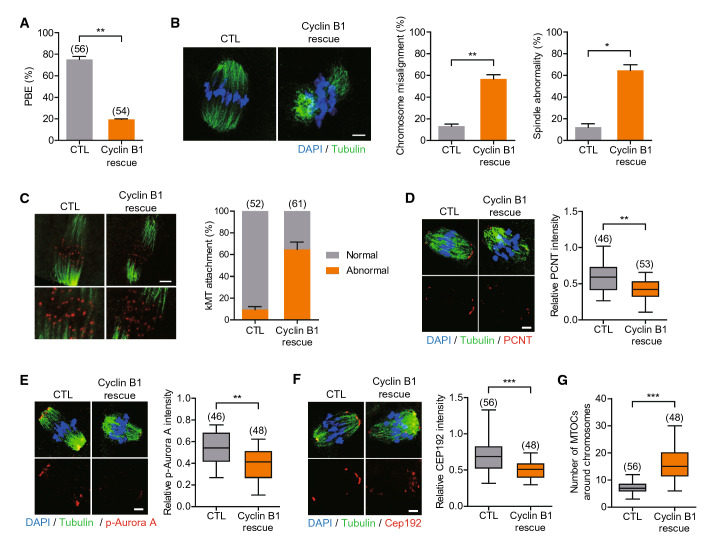
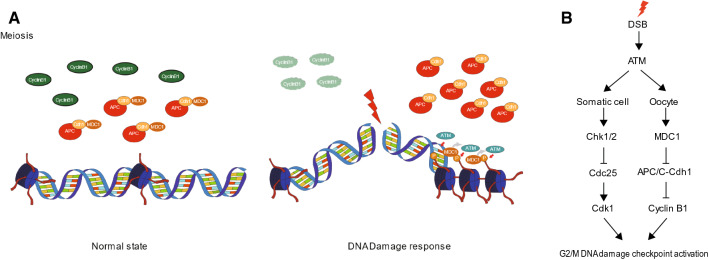
Mammalian oocytes are particularly susceptible to accumulating DNA damage. However, unlike mitotic cells in which DNA damage induces G2 arrest by activating the ATM-Chk1/2-Cdc25 pathway, oocytes readily enter M-phase immediately following DNA damage. This implies a lack of a robust canonical G2/M DNA damage checkpoint in oocytes. Here we show that MDC1 plays a non-canonical role in controlling G2/M transition by regulating APC/C-Cdh1-mediated cyclin B1 degradation in response to DNA damage in mouse oocytes. Depletion of MDC1 impaired M-phase entry by decreasing cyclin B1 levels via the APC/C-Cdh1 pathway. Notably, the APC/C-Cdh1 regulation mediated by MDC1 was achieved by a direct interaction between MDC1 and APC/C-Cdh1. This interaction was transiently disrupted after DNA damage with a concomitant increase in Cdh1 levels, which, in turn, decreased cyclin B1 levels and delayed M-phase entry. Moreover, MDC1 depletion impaired spindle assembly by decreasing the integrity of microtubule organizing centers (MTOCs). Therefore, our results demonstrate that MDC1 is an essential molecule in regulating G2/M transition in response to DNA damage and in regulating spindle assembly in mouse oocytes. These results provide new insights into the regulation of the G2/M DNA damage checkpoint and cell cycle control in oocytes.
Keywords: APC/C-Cdh1; DNA damage; G2/M transition; MDC1; Oocytes; Spindle assembly.
© 2022. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors have no conflict of interests to declare.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
