

Structure and Mechanism of the Lipid Flippase MurJ
- PMID: 35320686
- PMCID: PMC10108830
- DOI: 10.1146/annurev-biochem-040320-105145
Structure and Mechanism of the Lipid Flippase MurJ
Abstract
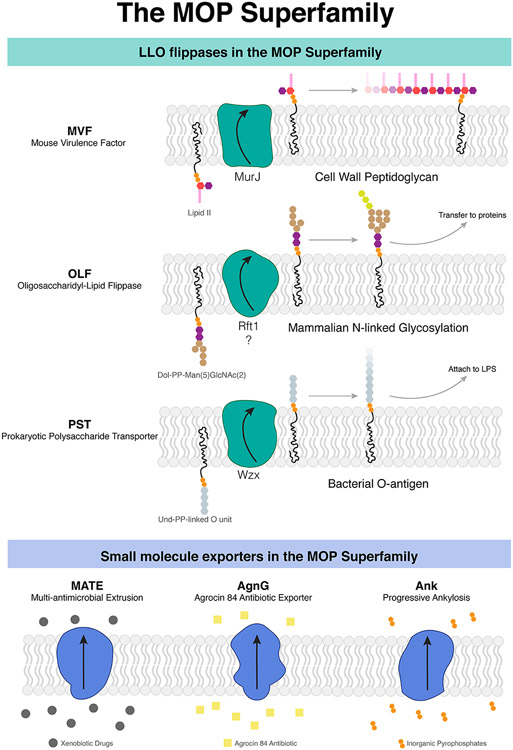
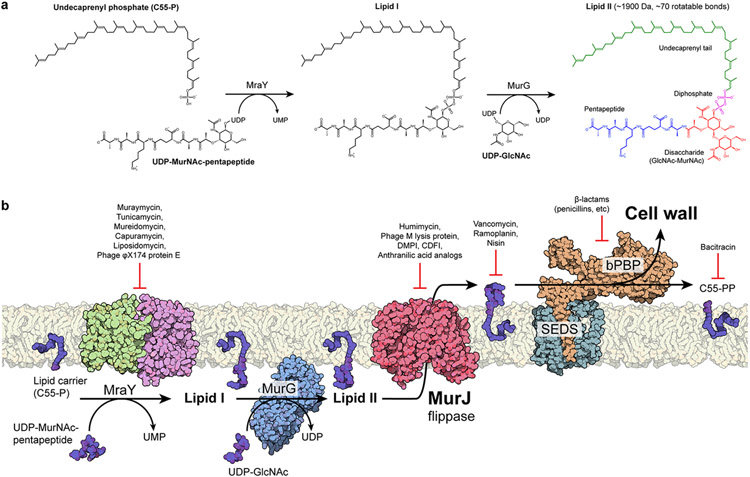
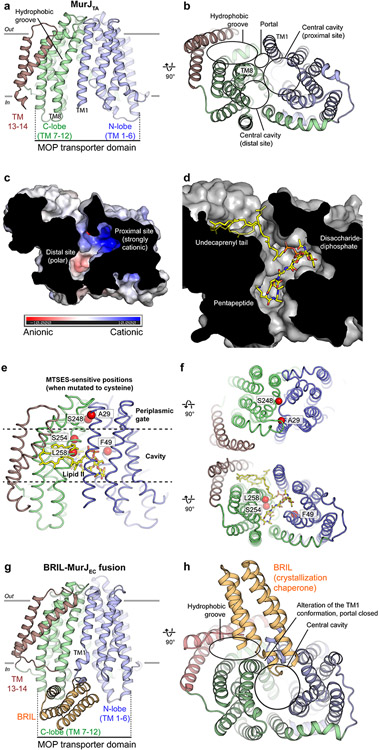
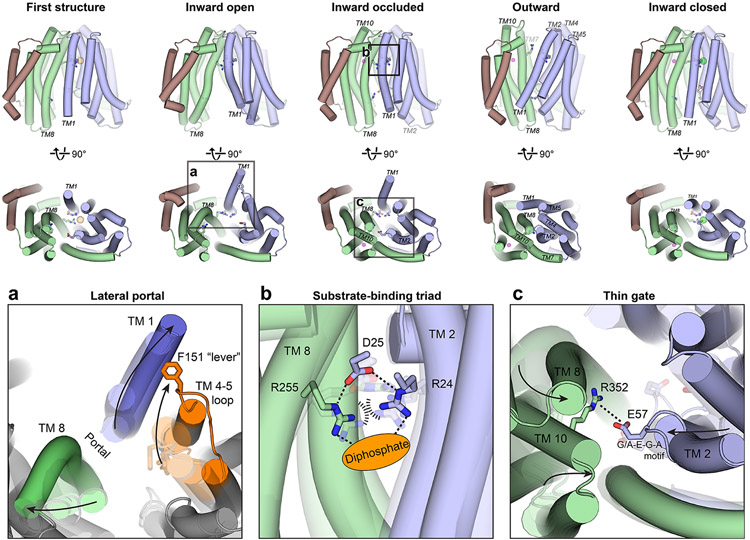
Biosynthesis of many important polysaccharides (including peptidoglycan, lipopolysaccharide, and N-linked glycans) necessitates the transport of lipid-linked oligosaccharides (LLO) across membranes from their cytosolic site of synthesis to their sites of utilization. Much of our current understanding of LLO transport comes from genetic, biochemical, and structural studies of the multidrug/oligosaccharidyl-lipid/polysaccharide (MOP) superfamily protein MurJ, which flips the peptidoglycan precursor lipid II. MurJ plays a pivotal role in bacterial cell wall synthesis and is an emerging antibiotic target. Here, we review the mechanism of LLO flipping by MurJ, including the structural basis for lipid II flipping and ion coupling. We then discuss inhibition of MurJ by antibacterials, including humimycins and the phage M lysis protein, as well as how studies on MurJ could provide insight into other flippases, both within and beyond the MOP superfamily.
Keywords: antibiotic; cell wall; lipid; peptidoglycan; transporter.
Figures
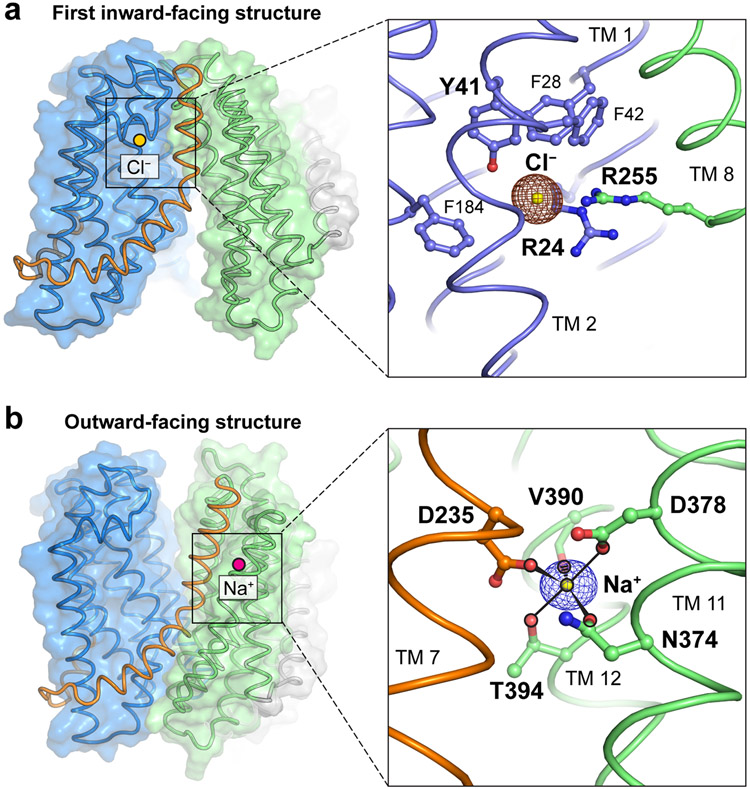
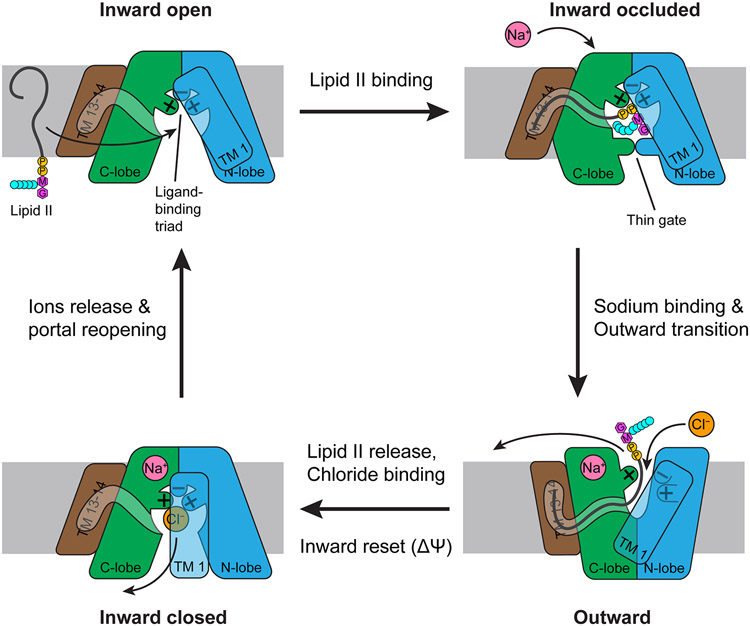
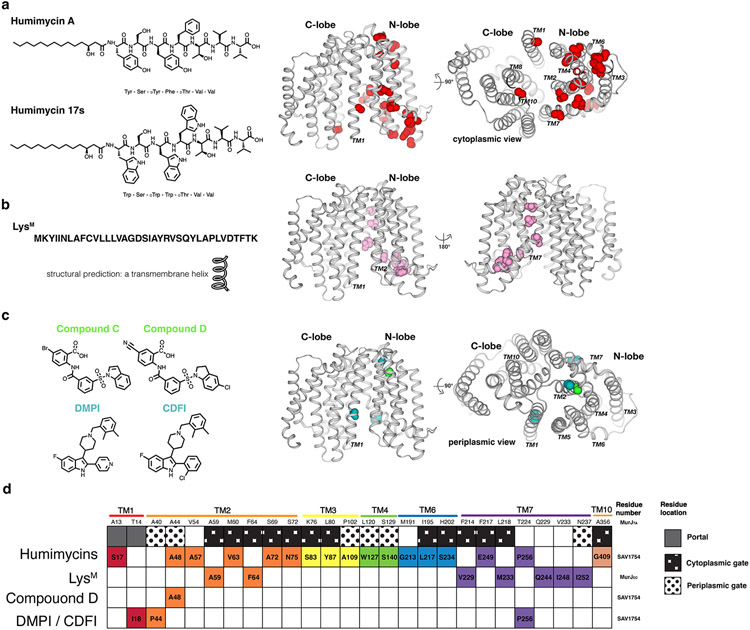
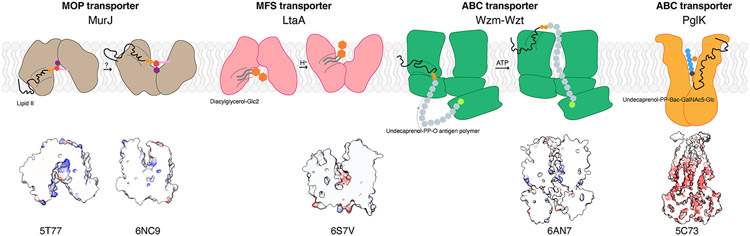
References
-
- Segawa K, Nagata S. 2015. An Apoptotic 'Eat Me' Signal: Phosphatidylserine Exposure. Trends Cell Biol 25: 639–50 - PubMed
-
- Henderson JC, Zimmerman SM, Crofts AA, Boll JM, Kuhns LG, et al. 2016. The Power of Asymmetry: Architecture and Assembly of the Gram-Negative Outer Membrane Lipid Bilayer. Annu Rev Microbiol 70: 255–78 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
