

Single-cell multiomics reveals persistence of HIV-1 in expanded cytotoxic T cell clones
- PMID: 35320704
- PMCID: PMC9203927
- DOI: 10.1016/j.immuni.2022.03.004
Single-cell multiomics reveals persistence of HIV-1 in expanded cytotoxic T cell clones
Abstract
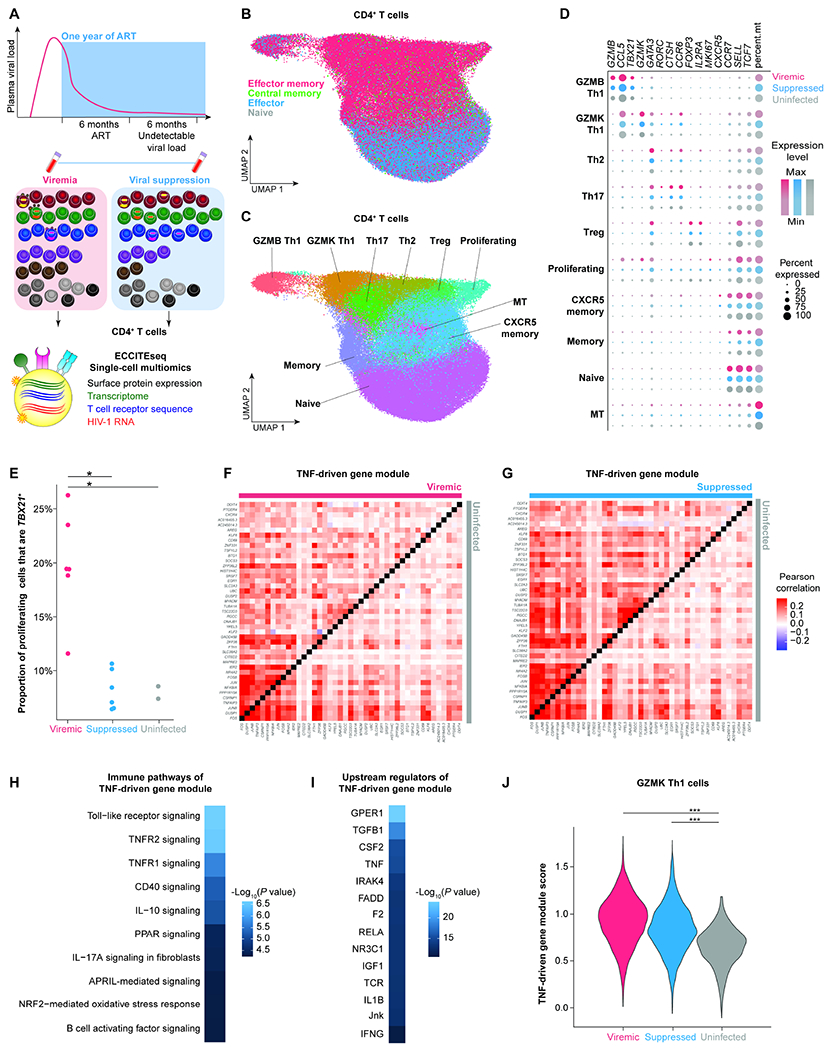
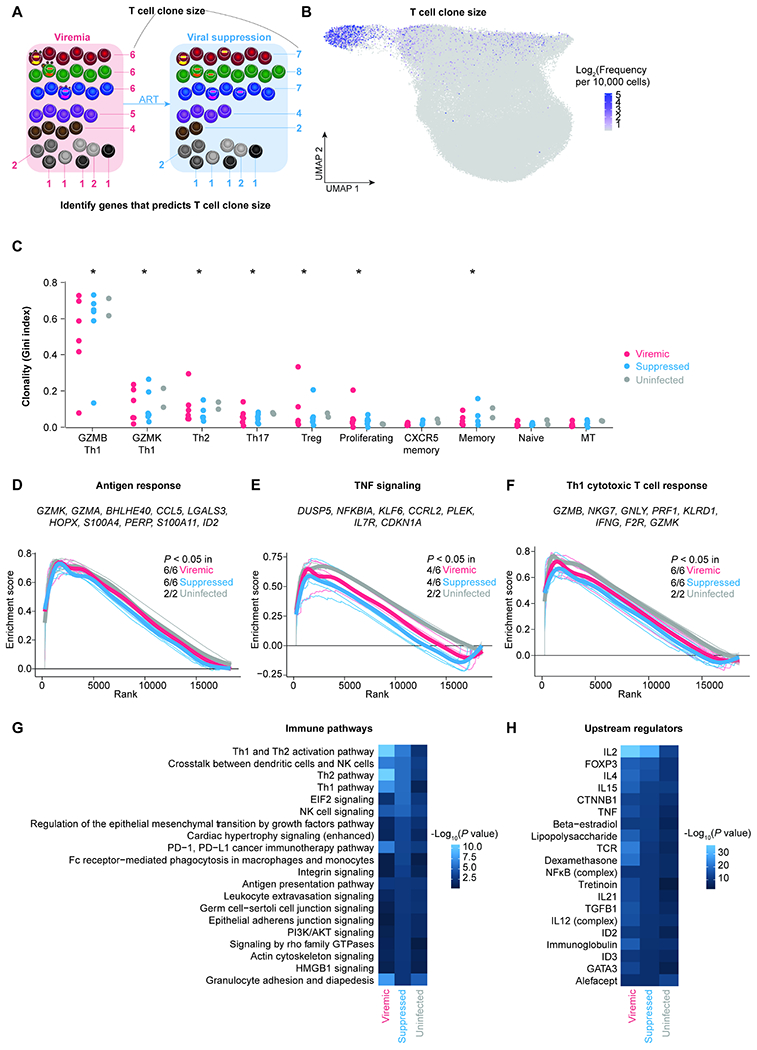
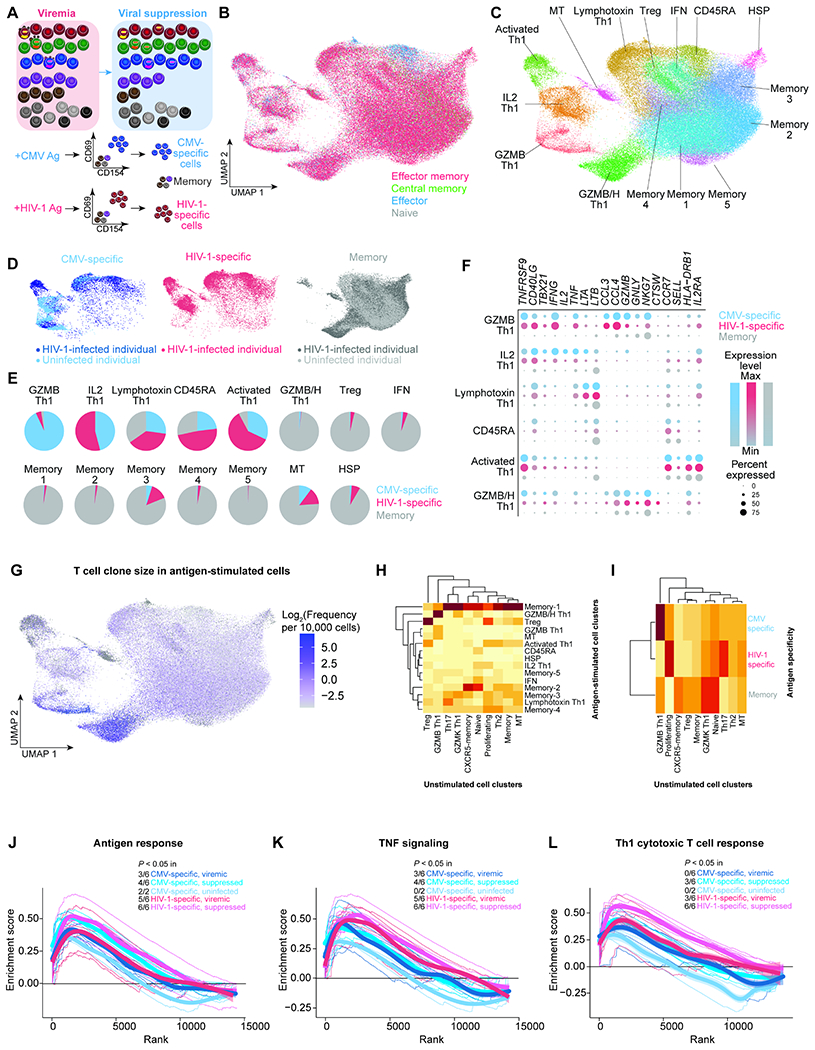
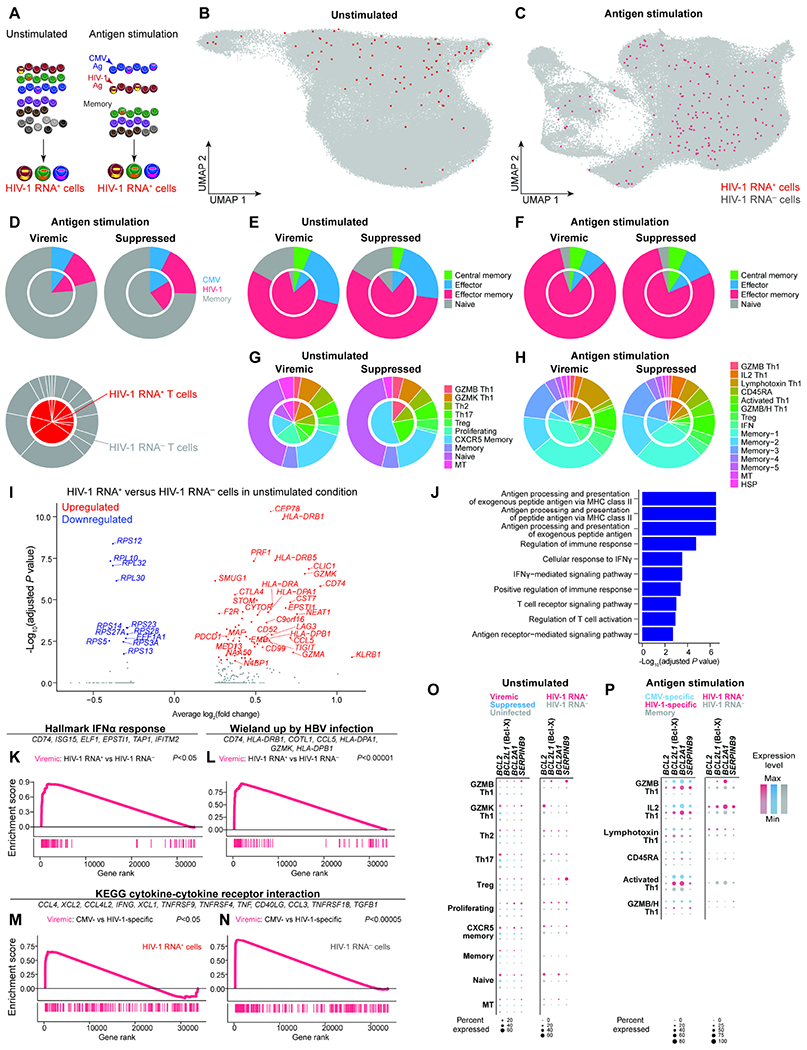
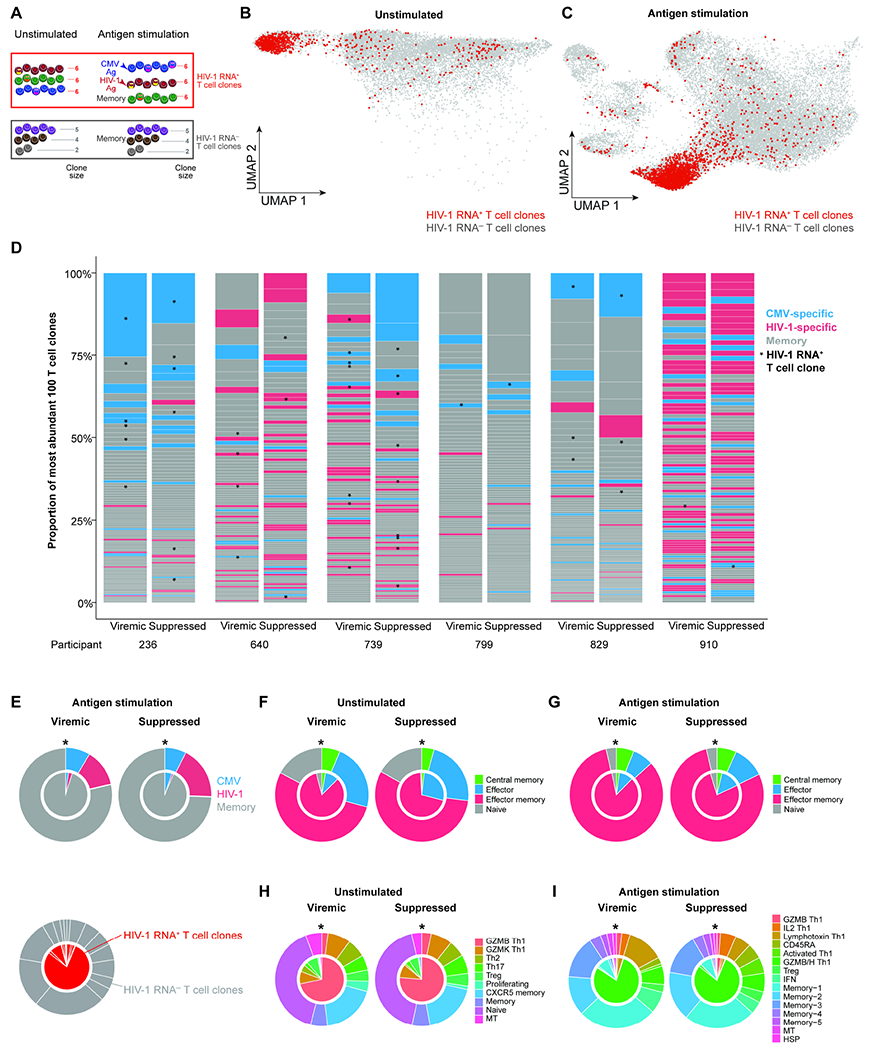
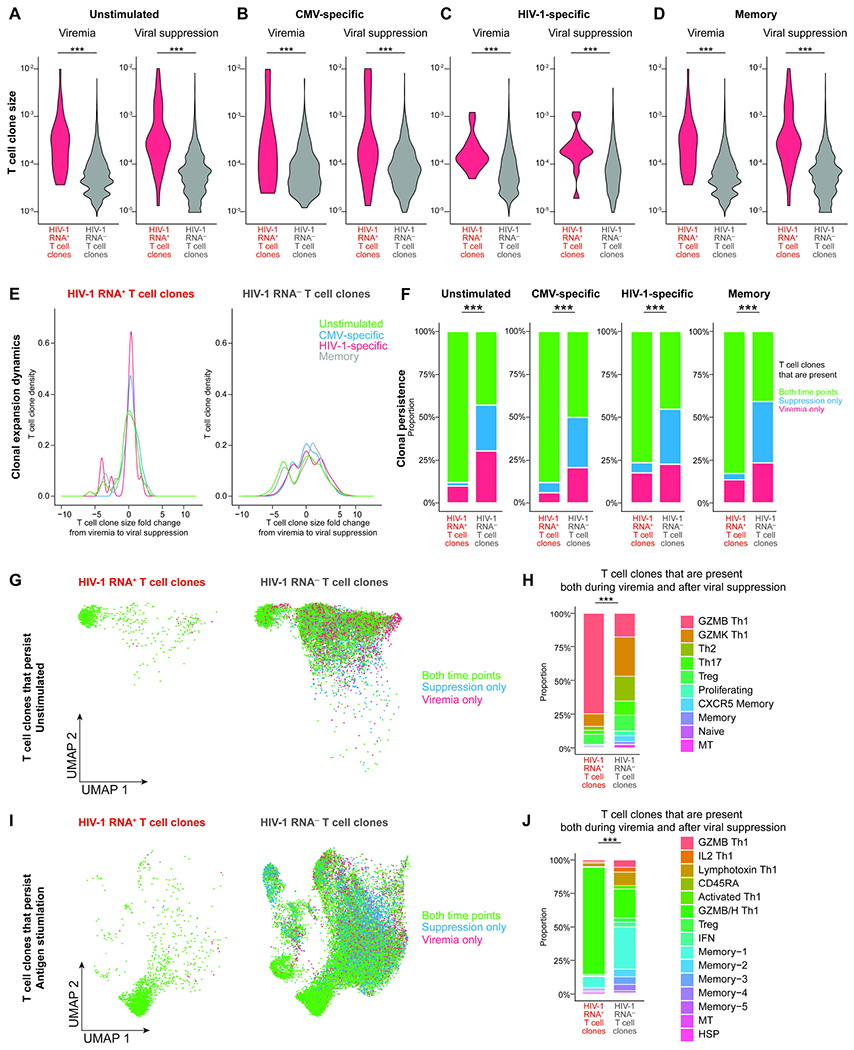
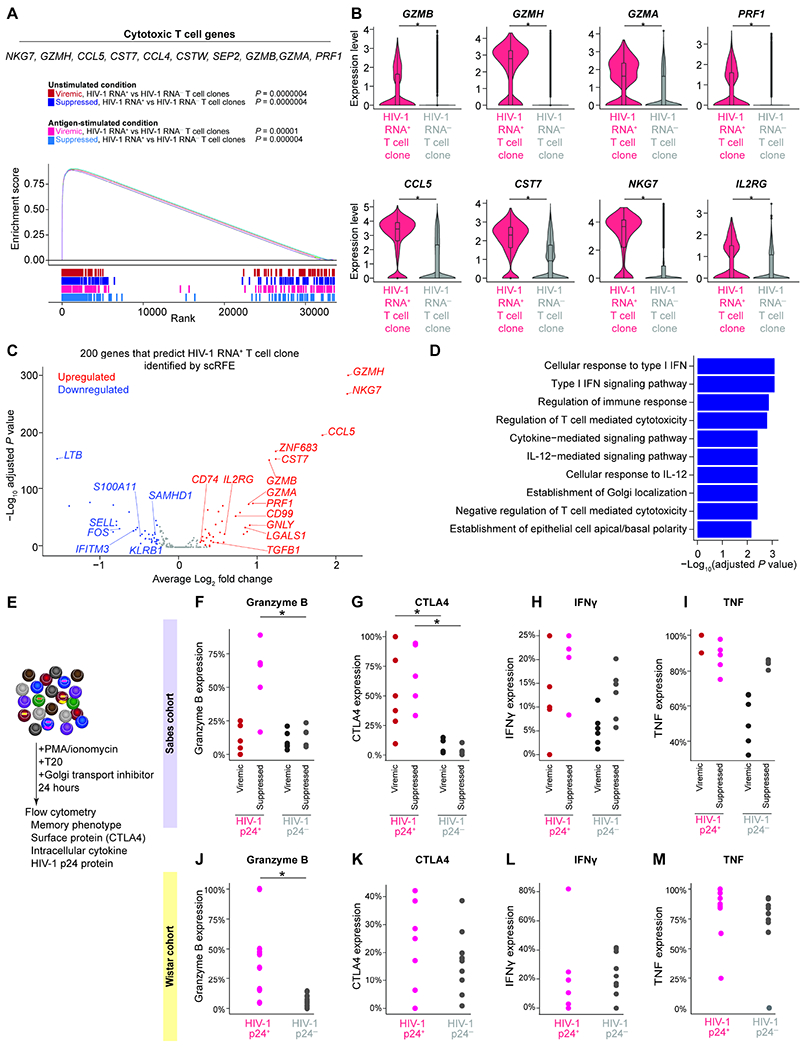
Understanding the drivers and markers of clonally expanding HIV-1-infected CD4+ T cells is essential for HIV-1 eradication. We used single-cell ECCITE-seq, which captures surface protein expression, cellular transcriptome, HIV-1 RNA, and TCR sequences within the same single cell to track clonal expansion dynamics in longitudinally archived samples from six HIV-1-infected individuals (during viremia and after suppressive antiretroviral therapy) and two uninfected individuals, in unstimulated conditions and after CMV and HIV-1 antigen stimulation. Despite antiretroviral therapy, persistent antigen and TNF responses shaped T cell clonal expansion. HIV-1 resided in Th1-polarized, antigen-responding T cells expressing BCL2 and SERPINB9 that may resist cell death. HIV-1 RNA+ T cell clones were larger in clone size, established during viremia, persistent after viral suppression, and enriched in GZMB+ cytotoxic effector memory Th1 cells. Targeting HIV-1-infected cytotoxic CD4+ T cells and drivers of clonal expansion provides another direction for HIV-1 eradication.
Keywords: HIV-1 latent reservoir; HIV-1 persistence; HIV-1-induced immune dysfunction; T cell expansion dynamics; TNF response; antigen stimulation; clonal expansion; cytotoxic CD4(+) T lymphocytes; granzyme B; single-cell RNA-seq.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Appay V, Zaunders JJ, Papagno L, Sutton J, Jaramillo A, Waters A, Easterbrook P, Grey P, Smith D, McMichael AJ, et al. (2002). Characterization of CD4(+) CTLs ex vivo. J Immunol 168, 5954–5958. - PubMed
-
- Bailey JR, Sedaghat AR, Kieffer T, Brennan T, Lee PK, Wind-Rotolo M, Haggerty CM, Kamireddi AR, Liu Y, Lee J, et al. (2006). Residual human immunodeficiency virus type 1 viremia in some patients on Antiretroviral therapy is dominated by a small number of invariant clones rarely found in circulating CD4(+) T cells. Journal of Virology 80, 6441–6457. - PMC - PubMed
-
- Barreiro LB, and Quintana-Murci L (2010). From evolutionary genetics to human immunology: how selection shapes host defence genes. Nat Rev Genet 11, 17–30. - PubMed
-
- Becht E, McInnes L, Healy J, Dutertre C-A, Kwok IWH, Ng LG, Ginhoux F, and Newell EW (2018). Dimensionality reduction for visualizing single-cell data using UMAP. Nature Biotechnology 37, 38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DA051906/DA/NIDA NIH HHS/United States
- UM1 AI164570/AI/NIAID NIH HHS/United States
- R01 AI145164/AI/NIAID NIH HHS/United States
- U01 DA053628/DA/NIDA NIH HHS/United States
- P50 AI150464/AI/NIAID NIH HHS/United States
- R01 DA032106/DA/NIDA NIH HHS/United States
- R01 DA040532/DA/NIDA NIH HHS/United States
- UM1 AI126620/AI/NIAID NIH HHS/United States
- R61 DA047037/DA/NIDA NIH HHS/United States
- T32 AI055403/AI/NIAID NIH HHS/United States
- UM1 AI164565/AI/NIAID NIH HHS/United States
- R35 GM143072/GM/NIGMS NIH HHS/United States
- UM1 DA051410/DA/NIDA NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- R33 DA047037/DA/NIDA NIH HHS/United States
- R01 MH125737/MH/NIMH NIH HHS/United States
- R01 AI141009/AI/NIAID NIH HHS/United States
- R37 AI147868/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
