

Telomerase deficiency reflects age-associated changes in CD4+ T cells
- PMID: 35321714
- PMCID: PMC8941756
- DOI: 10.1186/s12979-022-00273-0
Telomerase deficiency reflects age-associated changes in CD4+ T cells
Abstract
Background: Amongst other systemic changes, aging leads to an immune dysfunction. On the molecular level, a hallmark of aging is telomere shortening. The functional relevance of telomerase, an enzyme capable of elongating telomeres in T cells upon antigen stimulation, is not fully understood. Studying the impact of telomere shortening on CD4+ T cells and especially Th1 effector function can provide a better understanding on immune dysfunctions in elderly.
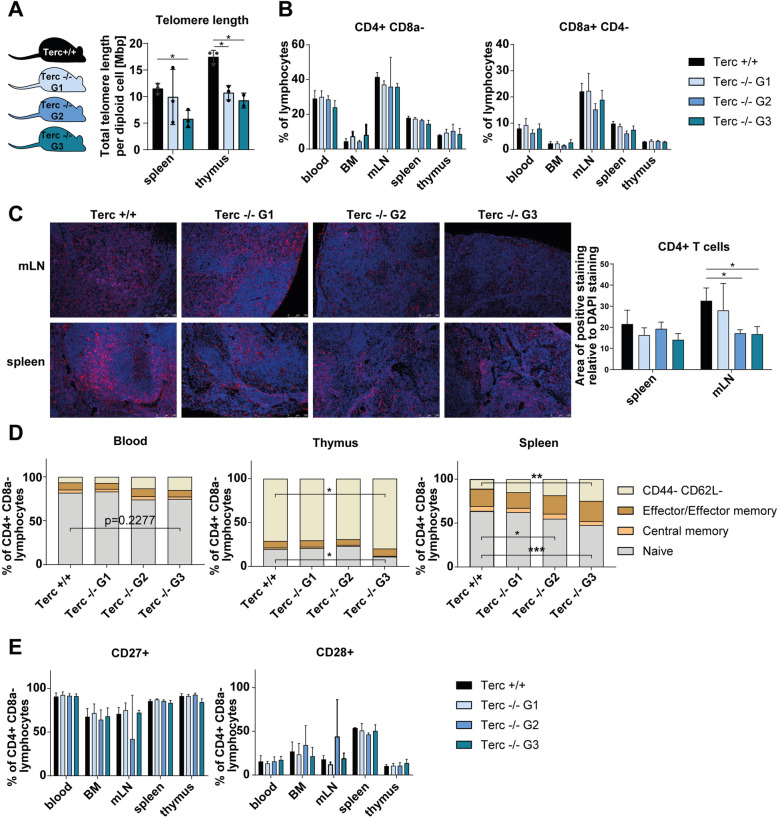
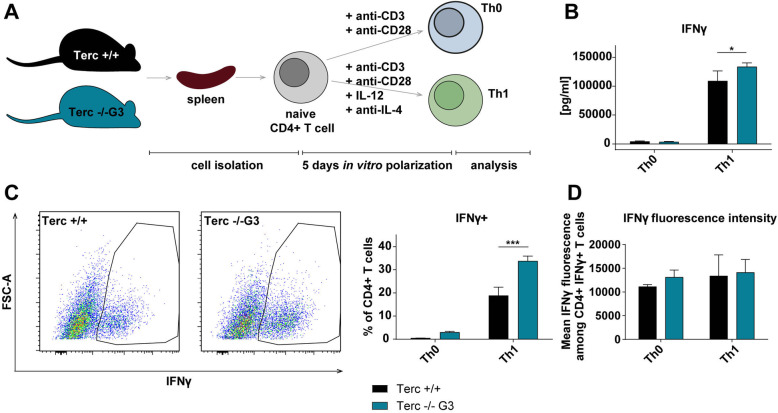
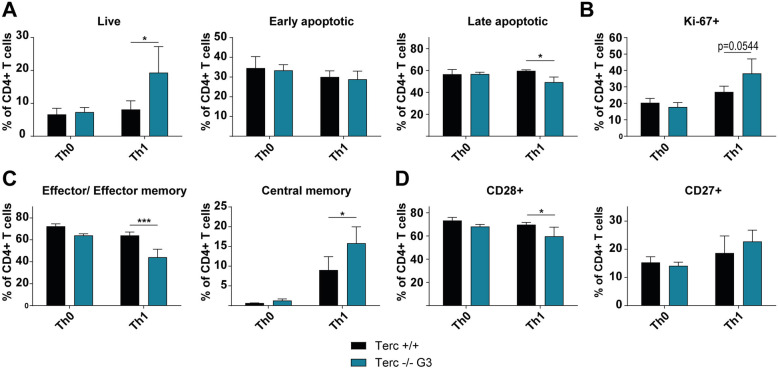
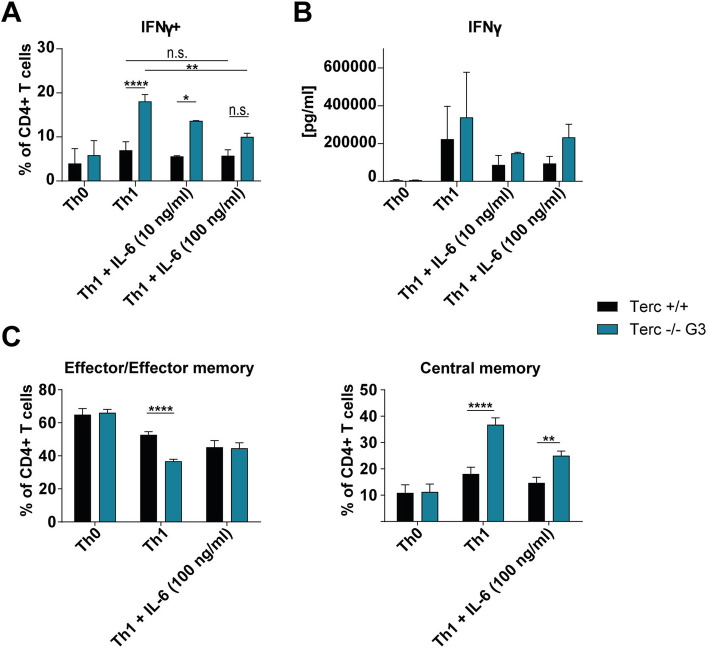
Results: We investigated T cell numbers and differentiation in telomerase-deficient (mTerc-/-) mice under steady-state conditions and the functional role of telomerase in CD4+ T cells using in vitro stimulation and Th1 polarization protocols by comparing T cells from mTerc-/- and control mice. We report reduced relative CD4+ T cell numbers in blood and secondary lymphoid organs and a relative decline in the naïve T cell population in thymus, blood and spleen of mTerc-/- mice compared to control mice. Importantly, after in vitro polarization, mTerc-/- G3 CD4+ T cells showed higher numbers of IFNγ-producing cells and reduced expression of CD28. Notably, telomerase-deficient T cells were more susceptible to inhibition of Th1 polarization by IL-6 in vitro. These results demonstrate that telomerase deficiency recapitulates several changes of CD4+ T cells seen in aged humans regarding the naïve T cell population, expression of CD28 and cytokine production.
Conclusion: Our data suggest that telomere shortening could play a key role in the aging of T cell immunity, with clinical implications for immune diseases and tumor development and that mTerc-/- mice are a suitable model to study aging-related defects of adaptive immunity.
Keywords: Aging; CD4-positive T-lymphocytes; Telomerase; Telomere shortening; Th1 cells.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials
