

Bioactivity and miRNome Profiling of Native Extracellular Vesicles in Human Induced Pluripotent Stem Cell-Cardiomyocyte Differentiation
- PMID: 35322574
- PMCID: PMC9130911
- DOI: 10.1002/advs.202104296
Bioactivity and miRNome Profiling of Native Extracellular Vesicles in Human Induced Pluripotent Stem Cell-Cardiomyocyte Differentiation
Abstract
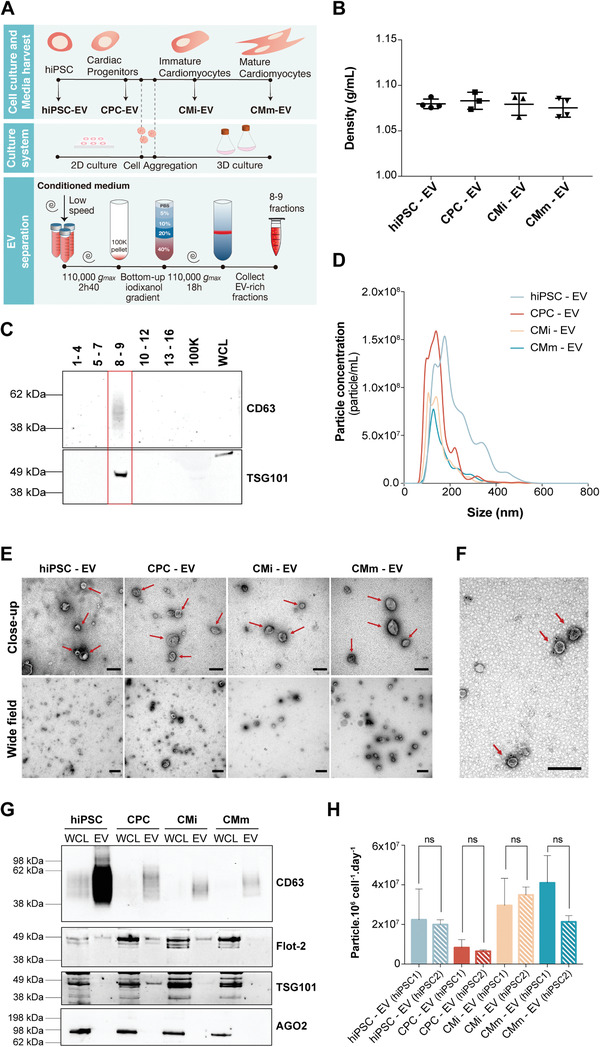
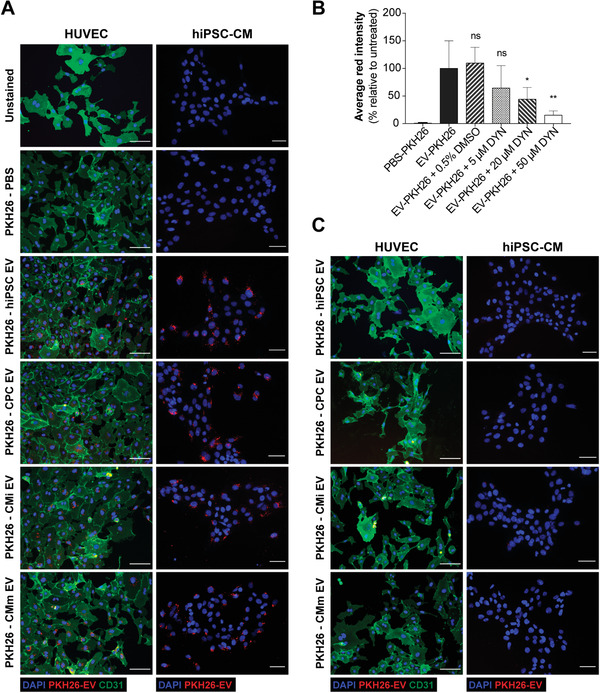
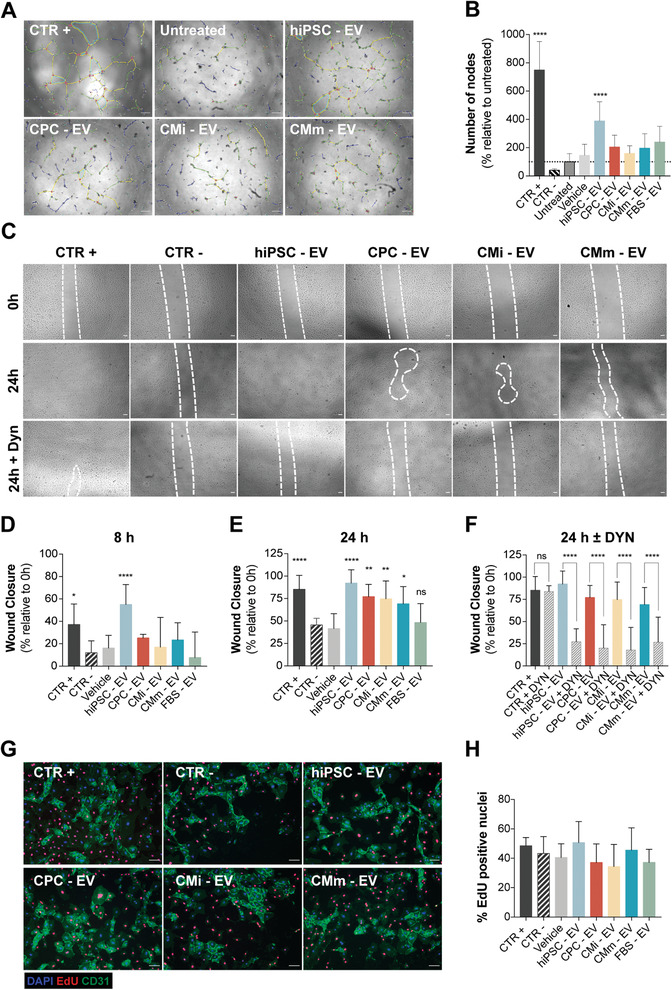
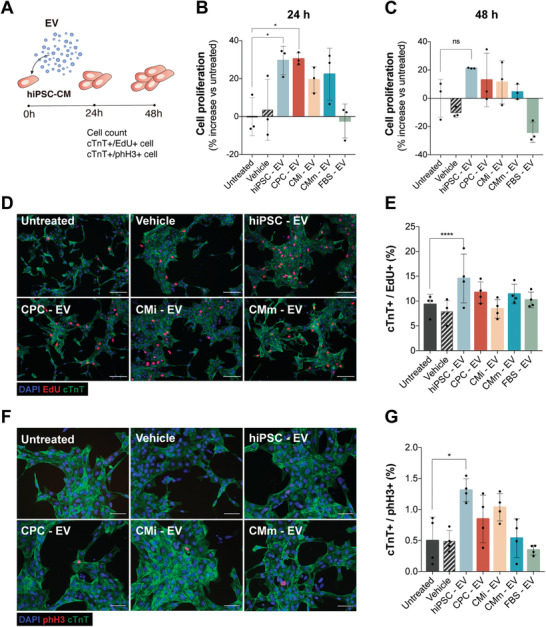
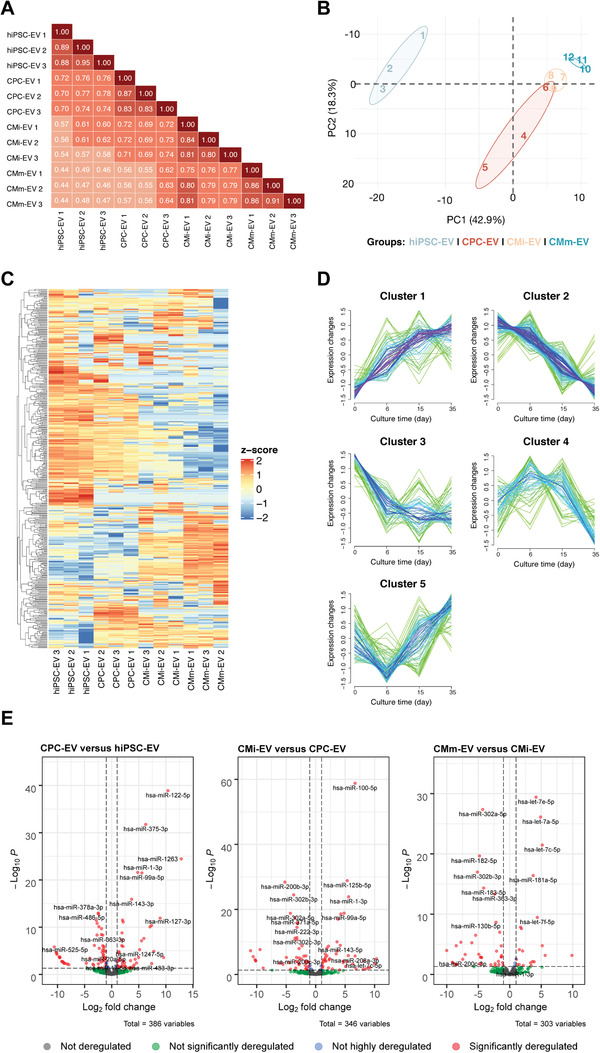
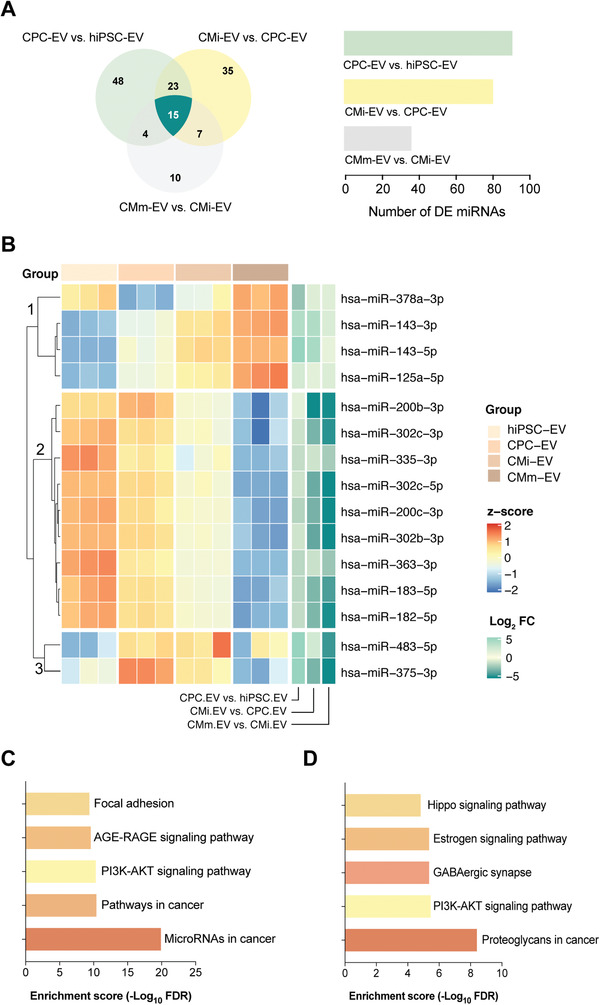
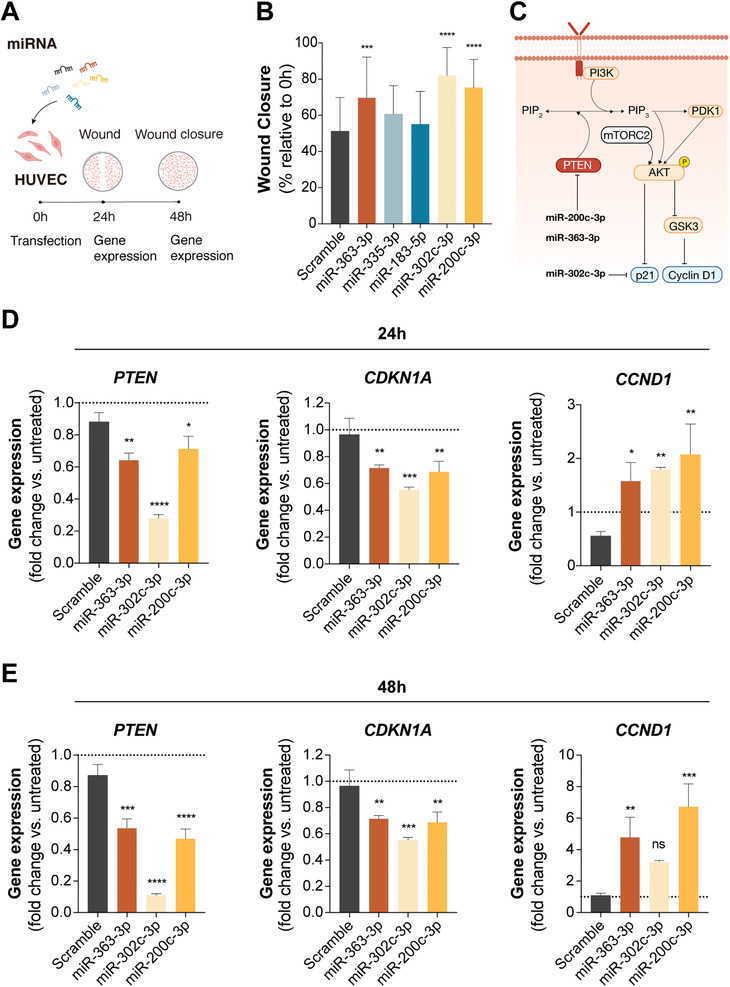
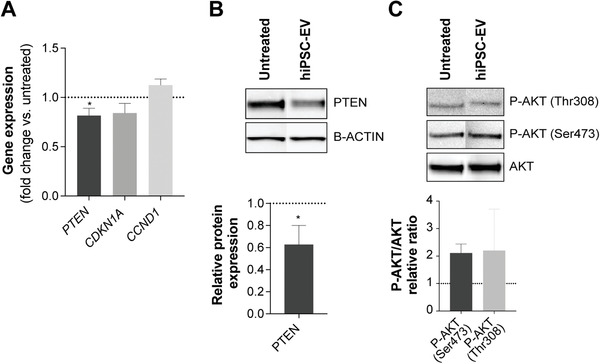
Extracellular vesicles (EV) are an attractive therapy to boost cardiac regeneration. Nevertheless, identification of native EV and corresponding cell platform(s) suitable for therapeutic application, is still a challenge. Here, EV are isolated from key stages of the human induced pluripotent stem cell-cardiomyocyte (hiPSC-CM) differentiation and maturation, i.e., from hiPSC (hiPSC-EV), cardiac progenitors, immature and mature cardiomyocytes, with the aim of identifying a promising cell biofactory for EV production, and pinpoint the genetic signatures of bioactive EV. EV secreted by hiPSC and cardiac derivatives show a typical size distribution profile and the expression of specific EV markers. Bioactivity assays show increased tube formation and migration in HUVEC treated with hiPSC-EV compared to EV from committed cell populations. hiPSC-EV also significantly increase cell cycle activity of hiPSC-CM. Global miRNA expression profiles, obtained by small RNA-seq analysis, corroborate an EV-miRNA pattern indicative of stem cell to cardiomyocyte specification, confirming that hiPSC-EV are enriched in pluripotency-associated miRNA with higher in vitro pro-angiogenic and pro-proliferative properties. In particular, a stemness maintenance miRNA cluster upregulated in hiPSC-EV targets the PTEN/PI3K/AKT pathway, involved in cell proliferation and survival. Overall, the findings validate hiPSC as cell biofactories for EV production for cardiac regenerative applications.
Keywords: cardiac regeneration; cell biofactory; extracellular vesicles; miRNome; small RNA-seq.
© 2022 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Expression of Extracellular Vesicle PIWI-Interacting RNAs Throughout hiPSC-Cardiomyocyte Differentiation.Front Physiol. 2022 Jun 16;13:926528. doi: 10.3389/fphys.2022.926528. eCollection 2022. Front Physiol. 2022. PMID: 35784878 Free PMC article.
-
Extracellular Vesicles from Human Induced Pluripotent Stem Cells Exhibit a Unique MicroRNA and CircRNA Signature.Int J Biol Sci. 2024 Nov 22;20(15):6255-6278. doi: 10.7150/ijbs.100113. eCollection 2024. Int J Biol Sci. 2024. PMID: 39664576 Free PMC article.
-
Extracellular vesicles from human cardiac stromal cells up-regulate cardiomyocyte protective responses to hypoxia.Stem Cell Res Ther. 2024 Oct 12;15(1):363. doi: 10.1186/s13287-024-03983-y. Stem Cell Res Ther. 2024. PMID: 39396003 Free PMC article.
-
Production of functional cardiomyocytes and cardiac tissue from human induced pluripotent stem cells for regenerative therapy.J Mol Cell Cardiol. 2022 Mar;164:83-91. doi: 10.1016/j.yjmcc.2021.11.008. Epub 2021 Nov 22. J Mol Cell Cardiol. 2022. PMID: 34822838 Review.
-
Extracellular vesicles and microRNAs in the regulation of cardiomyocyte differentiation and proliferation.Arch Biochem Biophys. 2023 Nov;749:109791. doi: 10.1016/j.abb.2023.109791. Epub 2023 Oct 18. Arch Biochem Biophys. 2023. PMID: 37858665 Review.
Cited by
-
A case for the study of native extracellular vesicles.Front Oncol. 2024 Jul 12;14:1430971. doi: 10.3389/fonc.2024.1430971. eCollection 2024. Front Oncol. 2024. PMID: 39091922 Free PMC article.
-
Expression of Extracellular Vesicle PIWI-Interacting RNAs Throughout hiPSC-Cardiomyocyte Differentiation.Front Physiol. 2022 Jun 16;13:926528. doi: 10.3389/fphys.2022.926528. eCollection 2022. Front Physiol. 2022. PMID: 35784878 Free PMC article.
-
Extracellular Vesicles from Human Induced Pluripotent Stem Cells Exhibit a Unique MicroRNA and CircRNA Signature.Int J Biol Sci. 2024 Nov 22;20(15):6255-6278. doi: 10.7150/ijbs.100113. eCollection 2024. Int J Biol Sci. 2024. PMID: 39664576 Free PMC article.
-
Clinically Expired Platelet Concentrates as a Source of Extracellular Vesicles for Targeted Anti-Cancer Drug Delivery.Pharmaceutics. 2023 Mar 15;15(3):953. doi: 10.3390/pharmaceutics15030953. Pharmaceutics. 2023. PMID: 36986815 Free PMC article.
-
Electric-Field-Driven Printed 3D Highly Ordered Microstructure with Cell Feature Size Promotes the Maturation of Engineered Cardiac Tissues.Adv Sci (Weinh). 2023 Apr;10(11):e2206264. doi: 10.1002/advs.202206264. Epub 2023 Feb 13. Adv Sci (Weinh). 2023. PMID: 36782337 Free PMC article.
References
-
- Yuana Y., Sturk A., Nieuwland R., Blood Rev. 2013, 27, 31. - PubMed
-
- Mathieu M., Martin‐Jaular L., Lavieu G., Théry C., Nat. Cell Biol. 2019, 21, 9. - PubMed
-
- Van Niel G., D'Angelo G., Raposo G., Nat. Rev. Mol. Cell Biol. 2018, 19, 213. - PubMed
-
- Théry C., Witwer K. W., Aikawa E., Alcaraz M. J., Anderson J. D., Andriantsitohaina R., Antoniou A., Arab T., Archer F., Atkin‐Smith G. K., Ayre D. C., Bach J. M., Bachurski D., Baharvand H., Balaj L., Baldacchino S., Bauer N. N., Baxter A. A., Bebawy M., Beckham C., Bedina Zavec A., Benmoussa A., Berardi A. C., Bergese P., Bielska E., Blenkiron C., Bobis‐Wozowicz S., Boilard E., Boireau W., Bongiovanni A., et al., J. Extracell. Vesicles 2018, 7, 1535750. - PMC - PubMed
-
- Sluijter J. P. G., Davidson S. M., Boulanger C. M., Buzás E. I., de Kleijn D. P. V., Engel F. B., Giricz Z., Hausenloy D. J., Kishore R., Lecour S., Leor J., Madonna R., Perrino C., Prunier F., Sahoo S., Schiffelers R. M., Schulz R., Van Laake L. W., Ytrehus K., Ferdinandy P., Cardiovasc. Res. 2018, 114, 19. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 874827/EU- H2020 Societal Challenges
- SAICTPAC/0047/2015/Fundação para a Ciência e Tecnologia
- PTDC/BTMSAL/32566/2017/Fundação para a Ciência e Tecnologia
- UIDB/04462/2020/Fundação para a Ciência e Tecnologia/Ministério da Ciência, Tecnologia e Ensino Superior
- UIDP/04462/2020/Fundação para a Ciência e Tecnologia/Ministério da Ciência, Tecnologia e Ensino Superior
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials