

Differential Selection on Caste-Associated Genes in a Subterranean Termite
- PMID: 35323522
- PMCID: PMC8955789
- DOI: 10.3390/insects13030224
Differential Selection on Caste-Associated Genes in a Subterranean Termite
Abstract
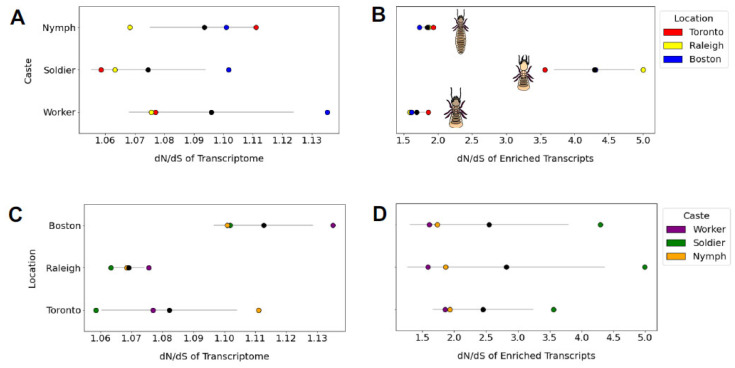
Analyzing the information-rich content of RNA can help uncover genetic events associated with social insect castes or other social polymorphisms. Here, we exploit a series of cDNA libraries previously derived from whole-body tissue of different castes as well as from three behaviourally distinct populations of the Eastern subterranean termite Reticulitermes flavipes. We found that the number (~0.5 M) of single nucleotide variants (SNVs) was roughly equal between nymph, worker and soldier caste libraries, but dN/dS (ratio of nonsynonymous to synonymous substitutions) analysis suggested that some of these variants confer a caste-specific advantage. Specifically, the dN/dS ratio was high (~4.3) for genes expressed in the defensively specialized soldier caste, relative to genes expressed by other castes (~1.7−1.8) and regardless of the North American population (Toronto, Raleigh, Boston) from which the castes were sampled. The populations, meanwhile, did show a large difference in SNV count but not in the manner expected from known demographic and behavioural differences; the highly invasive unicolonial population from Toronto was not the least diverse and did not show any other unique substitution patterns, suggesting any past bottleneck associated with invasion or with current unicoloniality has become obscured at the RNA level. Our study raises two important hypotheses relevant to termite sociobiology. First, the positive selection (dN/dS > 1) inferred for soldier-biased genes is presumably indirect and of the type mediated through kin selection, and second, the behavioural changes that accompany some social insect urban invasions (i.e., ‘unicoloniality’) may be detached from the loss-of-diversity expected from invasion bottlenecks.
Keywords: caste differentiation; kin selection; social insect invasions; urban entomology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Soldier-biased gene expression in a subterranean termite implies functional specialization of the defensive caste.Evol Dev. 2018 Jan;20(1):3-16. doi: 10.1111/ede.12243. Epub 2017 Nov 23. Evol Dev. 2018. PMID: 29168309
-
Comparative analysis of expressed sequence tags from three castes and two life stages of the termite Reticulitermes flavipes.BMC Genomics. 2010 Aug 6;11:463. doi: 10.1186/1471-2164-11-463. BMC Genomics. 2010. PMID: 20691076 Free PMC article.
-
Caste-biased patterns of brain investment in the subterranean termite Reticulitermes flavipes.iScience. 2024 May 22;27(6):110052. doi: 10.1016/j.isci.2024.110052. eCollection 2024 Jun 21. iScience. 2024. PMID: 38883809 Free PMC article.
-
The making of the defensive caste: Physiology, development, and evolution of the soldier differentiation in termites.Evol Dev. 2020 Nov;22(6):425-437. doi: 10.1111/ede.12335. Epub 2020 Apr 14. Evol Dev. 2020. PMID: 32291940 Review.
-
Developmental regulation of caste-specific characters in social-insect polyphenism.Evol Dev. 2005 Mar-Apr;7(2):122-9. doi: 10.1111/j.1525-142X.2005.05014.x. Evol Dev. 2005. PMID: 15733310 Review.
Cited by
-
Multi-Omic Analysis Reveals Population Differentiation and Signatures of Social Evolution in Tetragonula Stingless Bees.Mol Ecol. 2025 Jul;34(13):e17823. doi: 10.1111/mec.17823. Epub 2025 Jun 11. Mol Ecol. 2025. PMID: 40497295 Free PMC article.
References
-
- Pamilo P., Gertsch P., Thorén P., Seppä P. Molecular population genetics of social insects. Annu. Rev. Ecol. Syst. 1997;28:1–25. doi: 10.1146/annurev.ecolsys.28.1.1. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
