

The Cdkn2a gene product p19 alternative reading frame (p19ARF) is a critical regulator of IFNβ-mediated Lyme arthritis
- PMID: 35324997
- PMCID: PMC8946740
- DOI: 10.1371/journal.ppat.1010365
The Cdkn2a gene product p19 alternative reading frame (p19ARF) is a critical regulator of IFNβ-mediated Lyme arthritis
Abstract
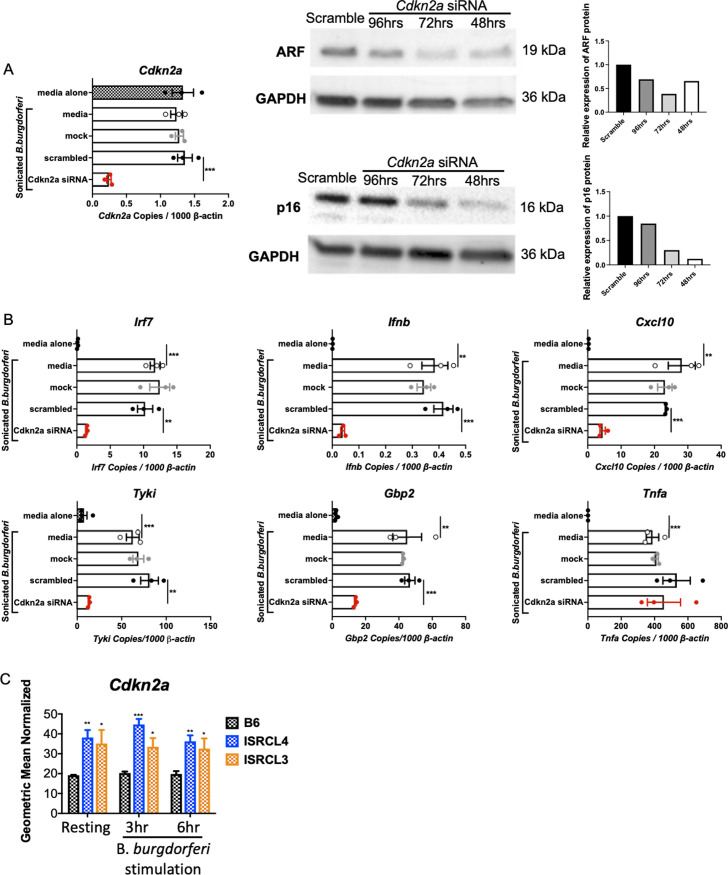
Type I interferon (IFN) has been identified in patients with Lyme disease, and its abundant expression in joint tissues of C3H mice precedes development of Lyme arthritis. Forward genetics using C3H mice with severe Lyme arthritis and C57BL/6 (B6) mice with mild Lyme arthritis identified the Borrelia burgdorferi arthritis-associated locus 1 (Bbaa1) on chromosome 4 (Chr4) as a regulator of B. burgdorferi-induced IFNβ expression and Lyme arthritis severity. B6 mice introgressed with the C3H allele for Bbaa1 (B6.C3-Bbaa1 mice) displayed increased severity of arthritis, which is initiated by myeloid lineage cells in joints. Using advanced congenic lines, the physical size of the Bbaa1 interval has been reduced to 2 Mbp, allowing for identification of potential genetic regulators. Small interfering RNA (siRNA)-mediated silencing identified Cdkn2a as the gene responsible for Bbaa1 allele-regulated induction of IFNβ and IFN-stimulated genes (ISGs) in bone marrow-derived macrophages (BMDMs). The Cdkn2a-encoded p19 alternative reading frame (p19ARF) protein regulates IFNβ induction in BMDMs as shown by siRNA silencing and overexpression of ARF. In vivo studies demonstrated that p19ARF contributes to joint-specific induction of IFNβ and arthritis severity in B. burgdorferi-infected mice. p19ARF regulates B. burgdorferi-induced IFNβ in BMDMs by stabilizing the tumor suppressor p53 and sequestering the transcriptional repressor BCL6. Our findings link p19ARF regulation of p53 and BCL6 to the severity of IFNβ-induced Lyme arthritis in vivo and indicate potential novel roles for p19ARF, p53, and BCL6 in Lyme disease and other IFN hyperproduction syndromes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures









Similar articles
-
Borrelia burgdorferi arthritis-associated locus Bbaa1 regulates Lyme arthritis and K/B×N serum transfer arthritis through intrinsic control of type I IFN production.J Immunol. 2014 Dec 15;193(12):6050-60. doi: 10.4049/jimmunol.1401746. Epub 2014 Nov 5. J Immunol. 2014. PMID: 25378596 Free PMC article.
-
Genetic Control of Lyme Arthritis by Borrelia burgdorferi Arthritis-Associated Locus 1 Is Dependent on Localized Differential Production of IFN-β and Requires Upregulation of Myostatin.J Immunol. 2017 Nov 15;199(10):3525-3534. doi: 10.4049/jimmunol.1701011. Epub 2017 Oct 6. J Immunol. 2017. PMID: 28986440 Free PMC article.
-
Interval-specific congenic lines reveal quantitative trait Loci with penetrant lyme arthritis phenotypes on chromosomes 5, 11, and 12.Infect Immun. 2009 Aug;77(8):3302-11. doi: 10.1128/IAI.00396-09. Epub 2009 Jun 1. Infect Immun. 2009. PMID: 19487472 Free PMC article.
-
Gene expression profiling provides insights into the pathways involved in inflammatory arthritis development: murine model of Lyme disease.Exp Mol Pathol. 2008 Aug;85(1):20-7. doi: 10.1016/j.yexmp.2008.03.004. Epub 2008 Apr 8. Exp Mol Pathol. 2008. PMID: 18462718 Free PMC article. Review.
-
Hamster and murine models of severe destructive Lyme arthritis.Clin Dev Immunol. 2012;2012:504215. doi: 10.1155/2012/504215. Epub 2012 Feb 22. Clin Dev Immunol. 2012. PMID: 22461836 Free PMC article. Review.
Cited by
-
Hitchhiker's Guide to Borrelia burgdorferi.J Bacteriol. 2024 Sep 19;206(9):e0011624. doi: 10.1128/jb.00116-24. Epub 2024 Aug 14. J Bacteriol. 2024. PMID: 39140751 Free PMC article. Review.
-
Endogenous Linear Plasmids lp28-4 and lp25 Are Required for Infectivity and Restriction Protection in the Lyme Disease Spirochete Borrelia mayonii.Infect Immun. 2023 Mar 15;91(3):e0006123. doi: 10.1128/iai.00061-23. Epub 2023 Feb 28. Infect Immun. 2023. PMID: 36853005 Free PMC article.
-
Association of Persistent Symptoms after Lyme Neuroborreliosis and Increased Levels of Interferon-α in Blood.Emerg Infect Dis. 2023 Jun;29(6):1091-1101. doi: 10.3201/eid2906.221685. Emerg Infect Dis. 2023. PMID: 37209716 Free PMC article. Review.
-
Impact of E. muris infection on B. burgdorferi-induced joint pathology in mice.Front Immunol. 2024 Aug 20;15:1430419. doi: 10.3389/fimmu.2024.1430419. eCollection 2024. Front Immunol. 2024. PMID: 39229265 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
