

Dysregulation of serum prolactin links the hypothalamus with female nociceptors to promote migraine
- PMID: 35325034
- PMCID: PMC9890468
- DOI: 10.1093/brain/awac104
Dysregulation of serum prolactin links the hypothalamus with female nociceptors to promote migraine
Abstract
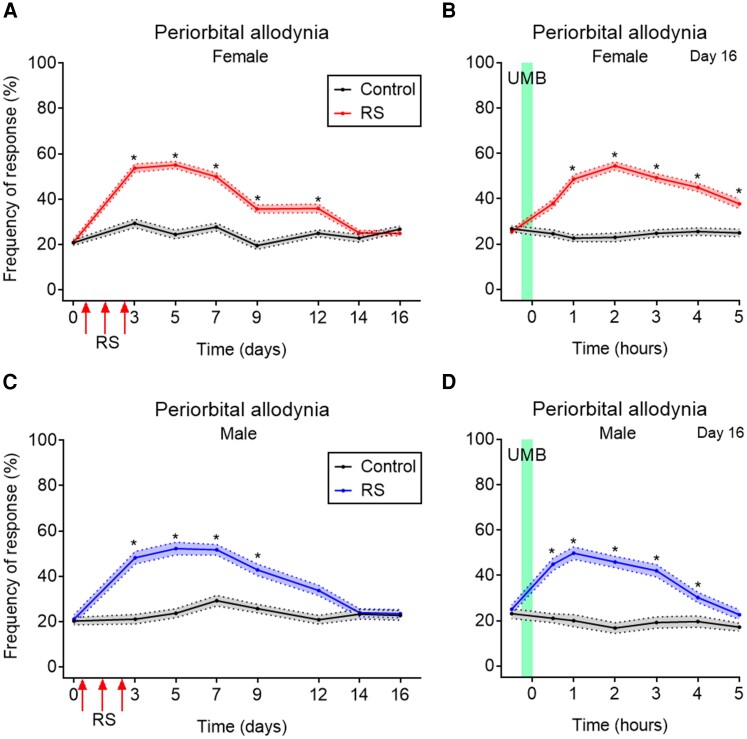
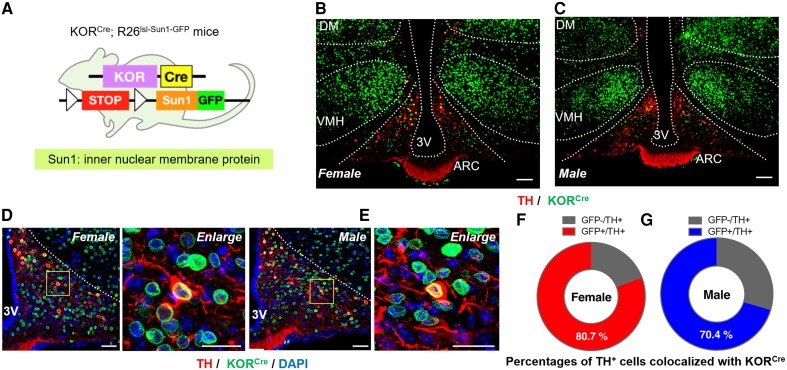
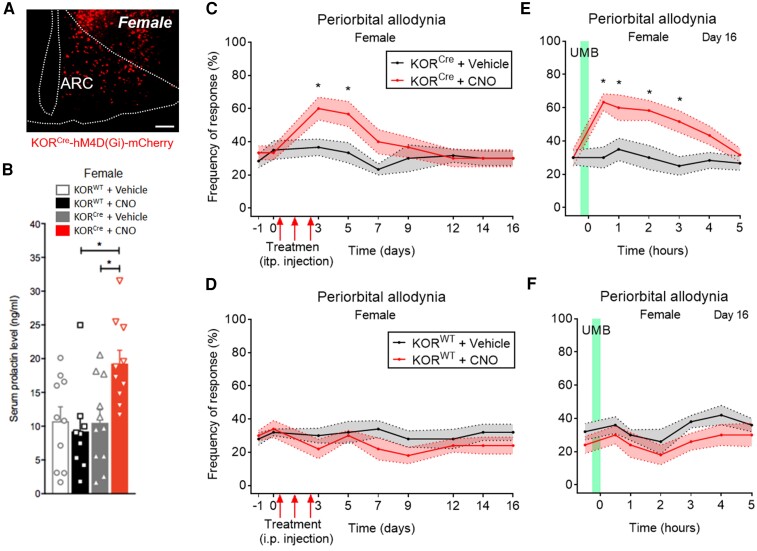
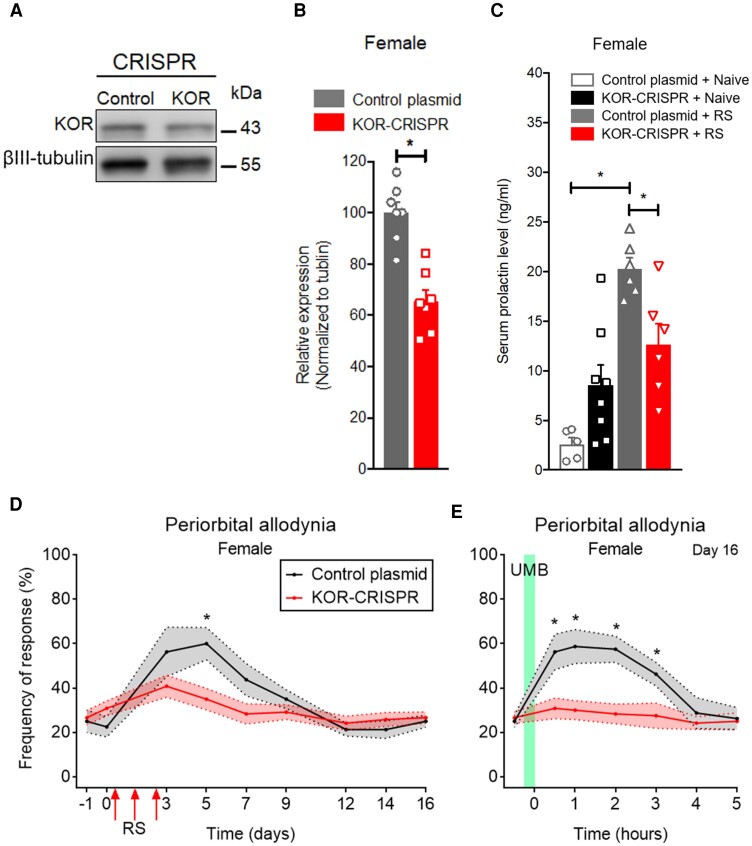
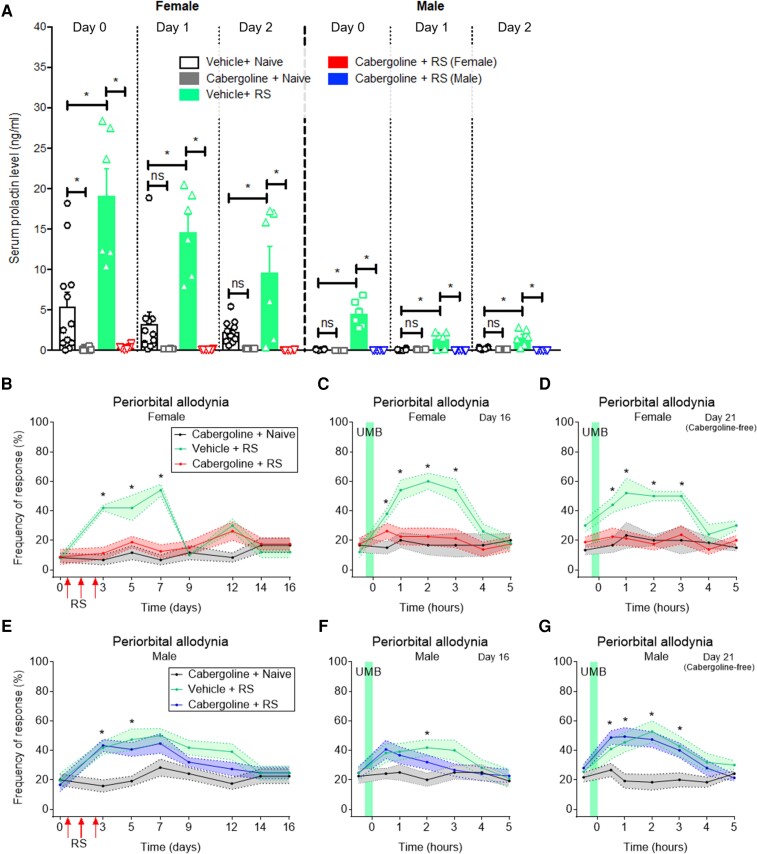
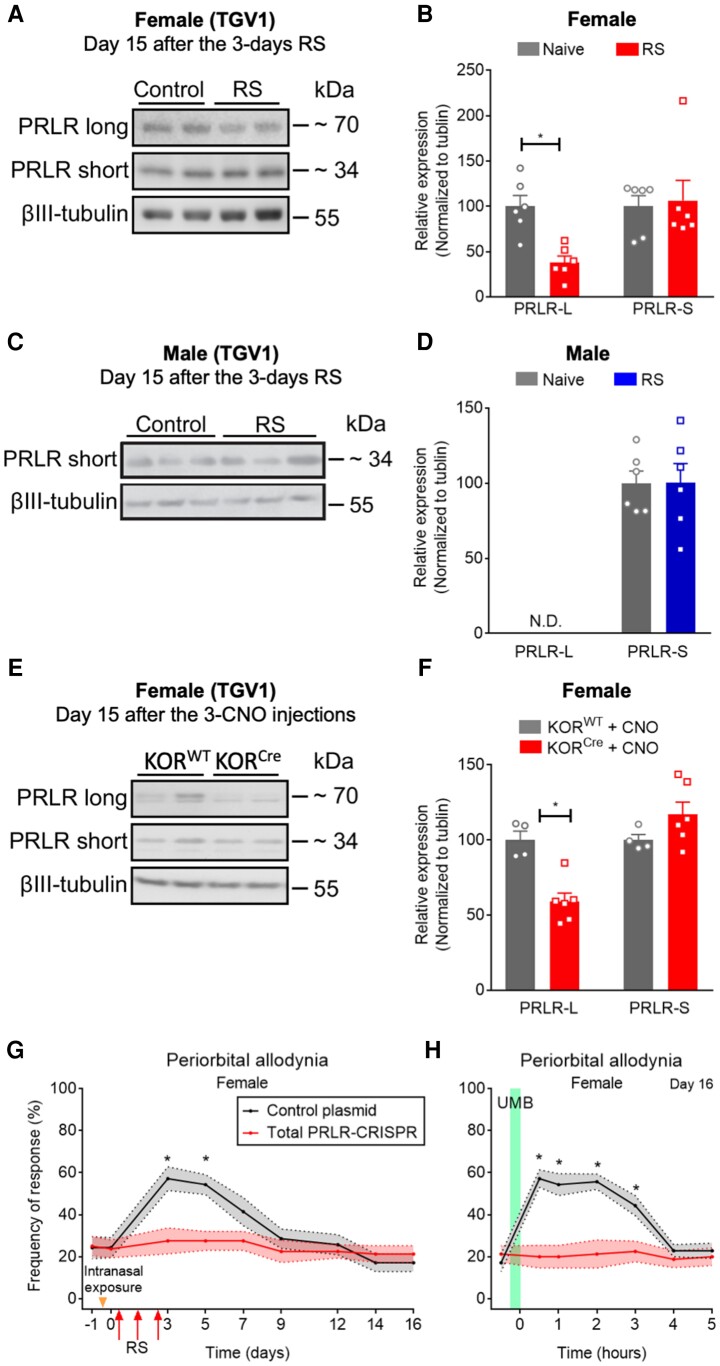
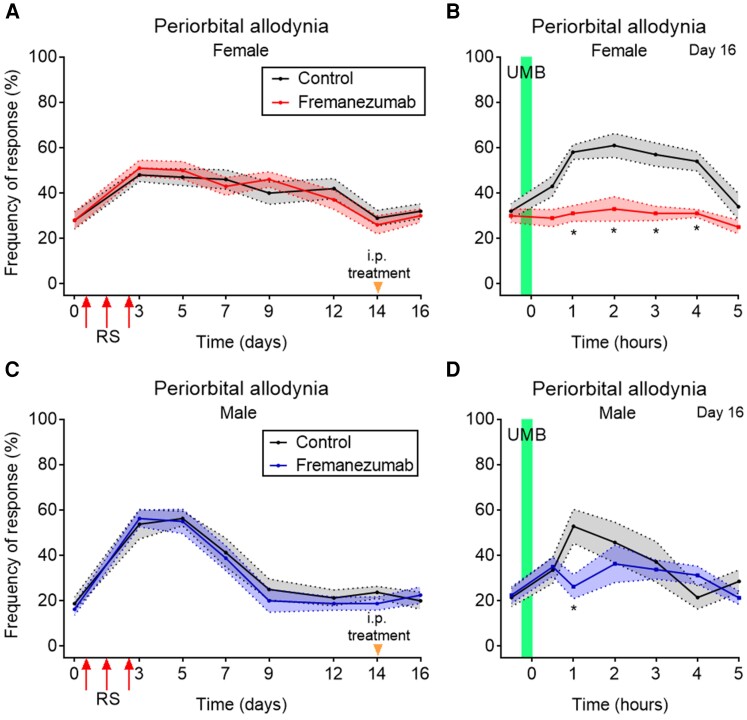
Migraine headache results from activation of meningeal nociceptors, however, the hypothalamus is activated many hours before the emergence of pain. How hypothalamic neural mechanisms may influence trigeminal nociceptor function remains unknown. Stress is a common migraine trigger that engages hypothalamic dynorphin/kappa opioid receptor (KOR) signalling and increases circulating prolactin. Prolactin acts at both long and short prolactin receptor isoforms that are expressed in trigeminal afferents. Following downregulation of the prolactin receptor long isoform, prolactin signalling at the prolactin receptor short isoform sensitizes nociceptors selectively in females. We hypothesized that stress may activate the kappa opioid receptor on tuberoinfundibular dopaminergic neurons to increase circulating prolactin leading to female-selective sensitization of trigeminal nociceptors through dysregulation of prolactin receptor isoforms. A mouse two-hit hyperalgesic priming model of migraine was used. Repeated restraint stress promoted vulnerability (i.e. first-hit priming) to a subsequent subthreshold (i.e. second-hit) stimulus from inhalational umbellulone, a TRPA1 agonist. Periorbital cutaneous allodynia served as a surrogate of migraine-like pain. Female and male KORCre; R26lsl-Sun1-GFP mice showed a high percentage of KORCre labelled neurons co-localized in tyrosine hydroxylase-positive cells in the hypothalamic arcuate nucleus. Restraint stress increased circulating prolactin to a greater degree in females. Stress-primed, but not control, mice of both sexes developed periorbital allodynia following inhalational umbellulone. Gi-DREADD activation (i.e. inhibition through Gi-coupled signalling) in KORCre neurons in the arcuate nucleus also increased circulating prolactin and repeated chemogenetic manipulation of these neurons primed mice of both sexes to umbellulone. Clustered regularly interspaced short palindromic repeats-Cas9 deletion of the arcuate nucleus KOR prevented restraint stress-induced prolactin release in female mice and priming from repeated stress episodes in both sexes. Inhibition of circulating prolactin occurred with systemic cabergoline, a dopamine D2 receptor agonist, blocked priming selectively in females. Repeated restraint stress downregulated the prolactin receptor long isoform in the trigeminal ganglia of female mice. Deletion of prolactin receptor in trigeminal ganglia by nasal clustered regularly interspaced short palindromic repeats-Cas9 targeting both prolactin receptor isoforms prevented stress-induced priming in female mice. Stress-induced activation of hypothalamic KOR increases circulating prolactin resulting in trigeminal downregulation of prolactin receptor long and pain responses to a normally innocuous TRPA1 stimulus. These are the first data that provide a mechanistic link between stress-induced hypothalamic activation and the trigeminal nociceptor effectors that produce trigeminal sensitization and migraine-like pain. This sexually dimorphic mechanism may help to explain female prevalence of migraine. KOR antagonists, currently in phase II clinical trials, may be useful as migraine preventives in both sexes, while dopamine agonists and prolactin/ prolactin receptor antibodies may improve therapy for migraine, and other stress-related neurological disorders, in females.
Keywords: hypothalamus; kappa opioid receptors; migraine; prolactin; stress.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Bourke JH, Langford RM, White PD. The common link between functional somatic syndromes may be central sensitisation. J Psychosom Res. 2015;78:228–236. - PubMed
-
- Nahman-Averbuch H, Granovsky Y, Coghill RC, et al. Waning of “conditioned pain modulation”: A novel expression of subtle pronociception in migraine. Headache. 2013;53:1104–1115. - PubMed
-
- Dodick DW. Migraine. Lancet. 2018;391:1315–1330. - PubMed
-
- Goadsby PJ, Holland PR. An update: pathophysiology of migraine. Neurol Clin. 2019;37:651–671. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
