

T lymphocytes as dynamic regulators of glioma pathobiology
- PMID: 35325210
- PMCID: PMC9527522
- DOI: 10.1093/neuonc/noac055
T lymphocytes as dynamic regulators of glioma pathobiology
Abstract
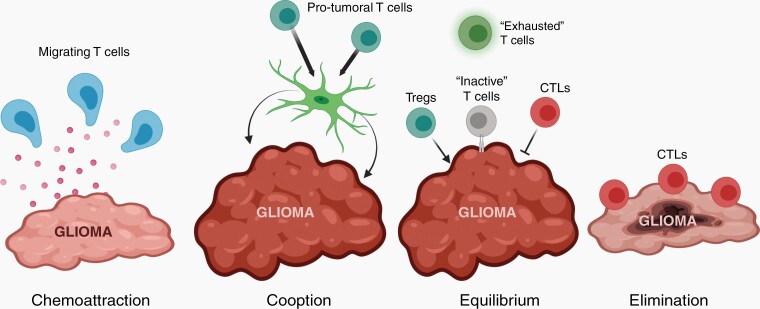
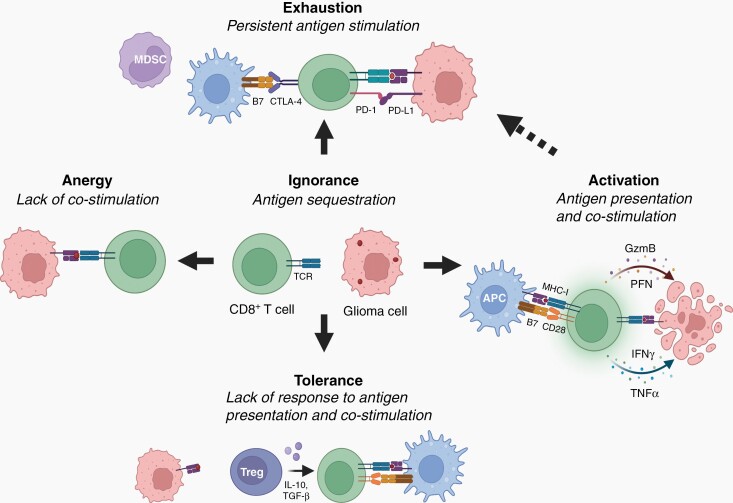
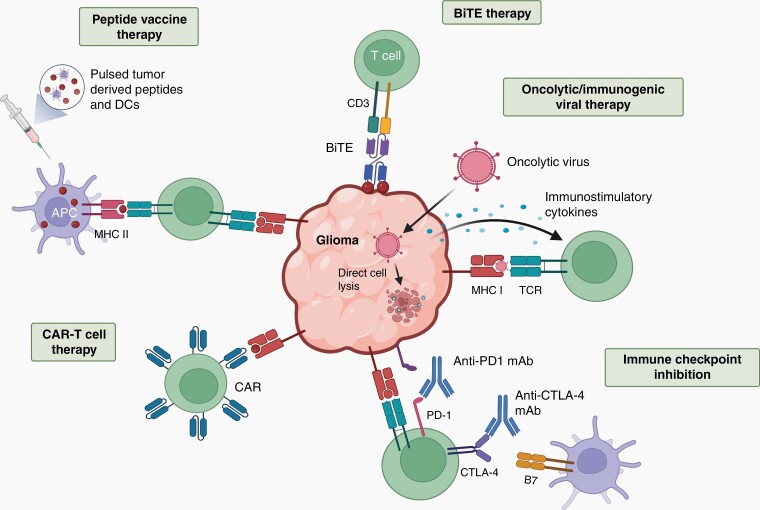
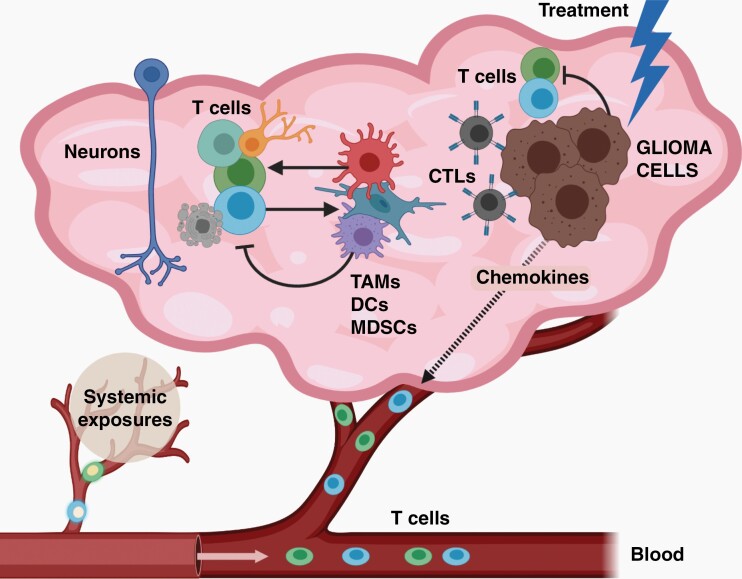
The brain tumor microenvironment contains numerous distinct types of nonneoplastic cells, which each serve a diverse set of roles relevant to the formation, maintenance, and progression of these central nervous system cancers. While varying in frequencies, monocytes (macrophages, microglia, and myeloid-derived suppressor cells), dendritic cells, natural killer cells, and T lymphocytes represent the most common nonneoplastic cellular constituents in low- and high-grade gliomas (astrocytomas). Although T cells are conventionally thought to target and eliminate neoplastic cells, T cells also exist in other states, characterized by tolerance, ignorance, anergy, and exhaustion. In addition, T cells can function as drivers of brain cancer growth, especially in low-grade gliomas. Since T cells originate in the blood and bone marrow sinuses, their capacity to function as both positive and negative regulators of glioma growth has ignited renewed interest in their deployment as immunotherapeutic agents. In this review, we discuss the roles of T cells in low- and high-grade glioma formation and progression, as well as the potential uses of modified T lymphocytes for brain cancer therapeutics.
Keywords: T cells; astrocytoma; glioblastoma; gliomagenesis; microglia; pediatric low-grade glioma; tumor microenvironment; tumor-associated monocytes.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Comment in
-
Understanding the relationship between gliomas and T cells: Paving the way for effective immunotherapy.Neuro Oncol. 2022 Oct 3;24(10):1658-1659. doi: 10.1093/neuonc/noac152. Neuro Oncol. 2022. PMID: 35660928 Free PMC article. No abstract available.
References
-
- Friebel E, Kapolou K, Unger S, et al. Single-cell mapping of human brain cancer reveals tumor-specific instruction of tissue-invading leukocytes. Cell. 2020; 181(7):1626–1642.e20. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
