

Facile discovery of surrogate cytokine agonists
- PMID: 35325595
- PMCID: PMC9021867
- DOI: 10.1016/j.cell.2022.02.025
Facile discovery of surrogate cytokine agonists
Abstract
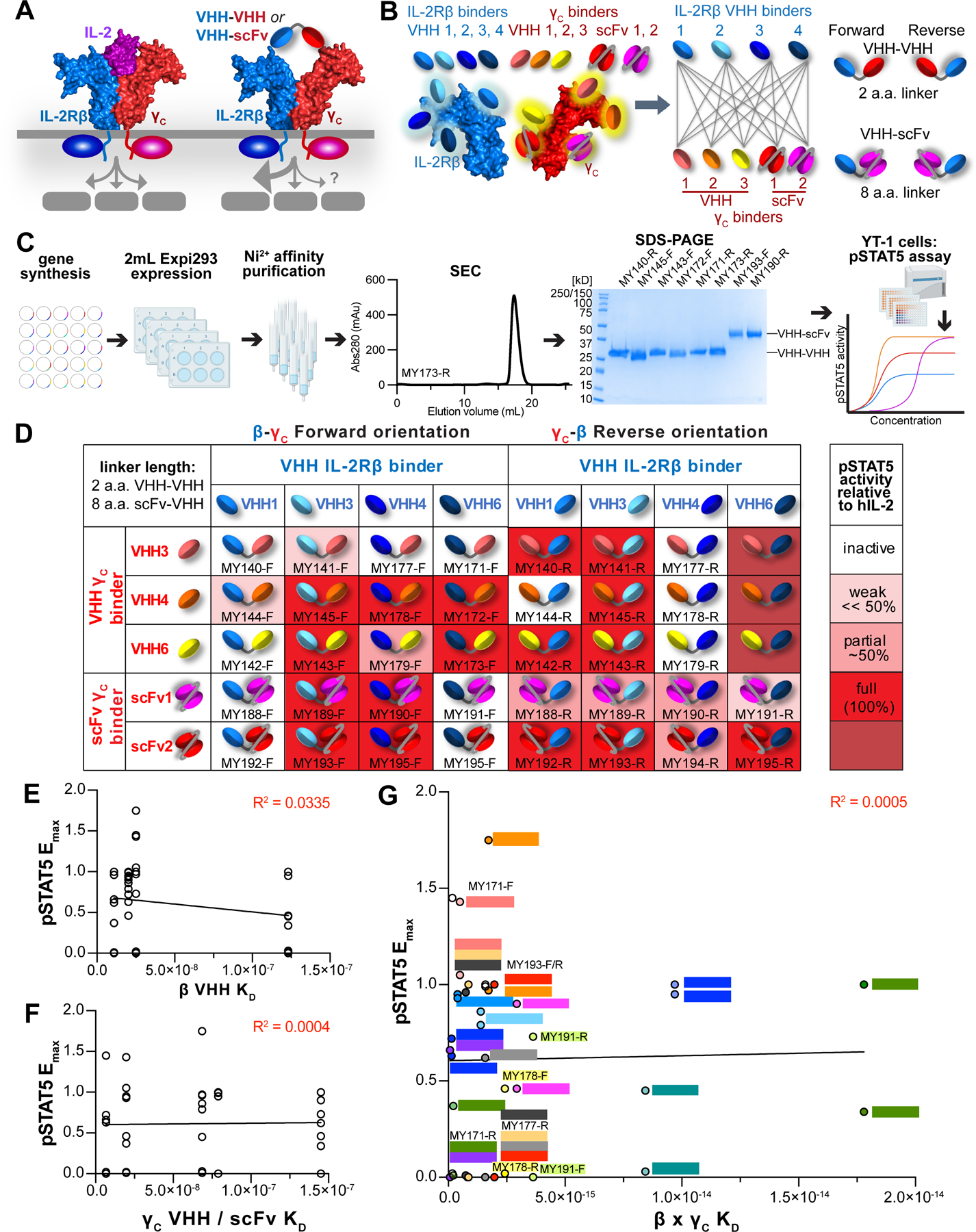
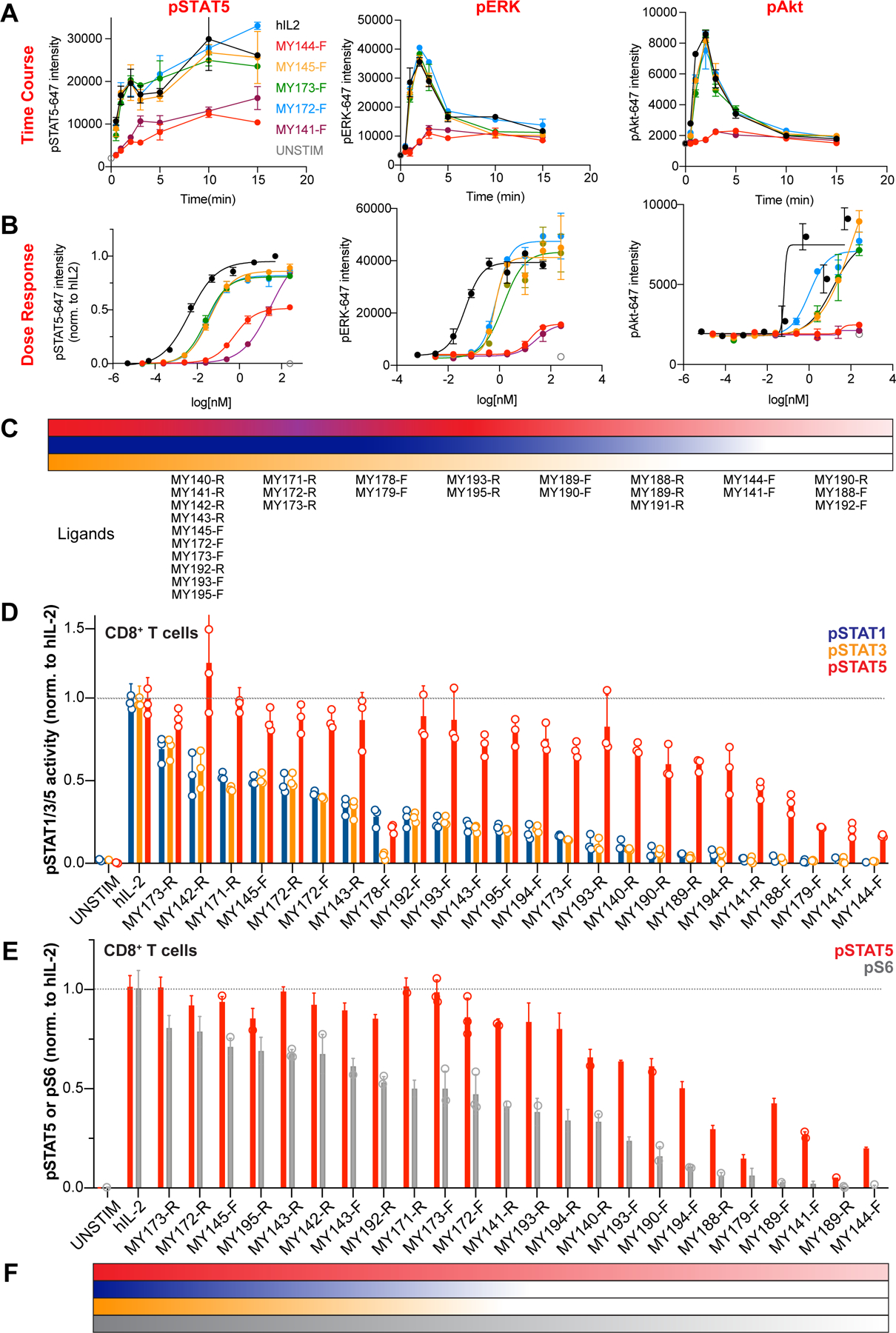
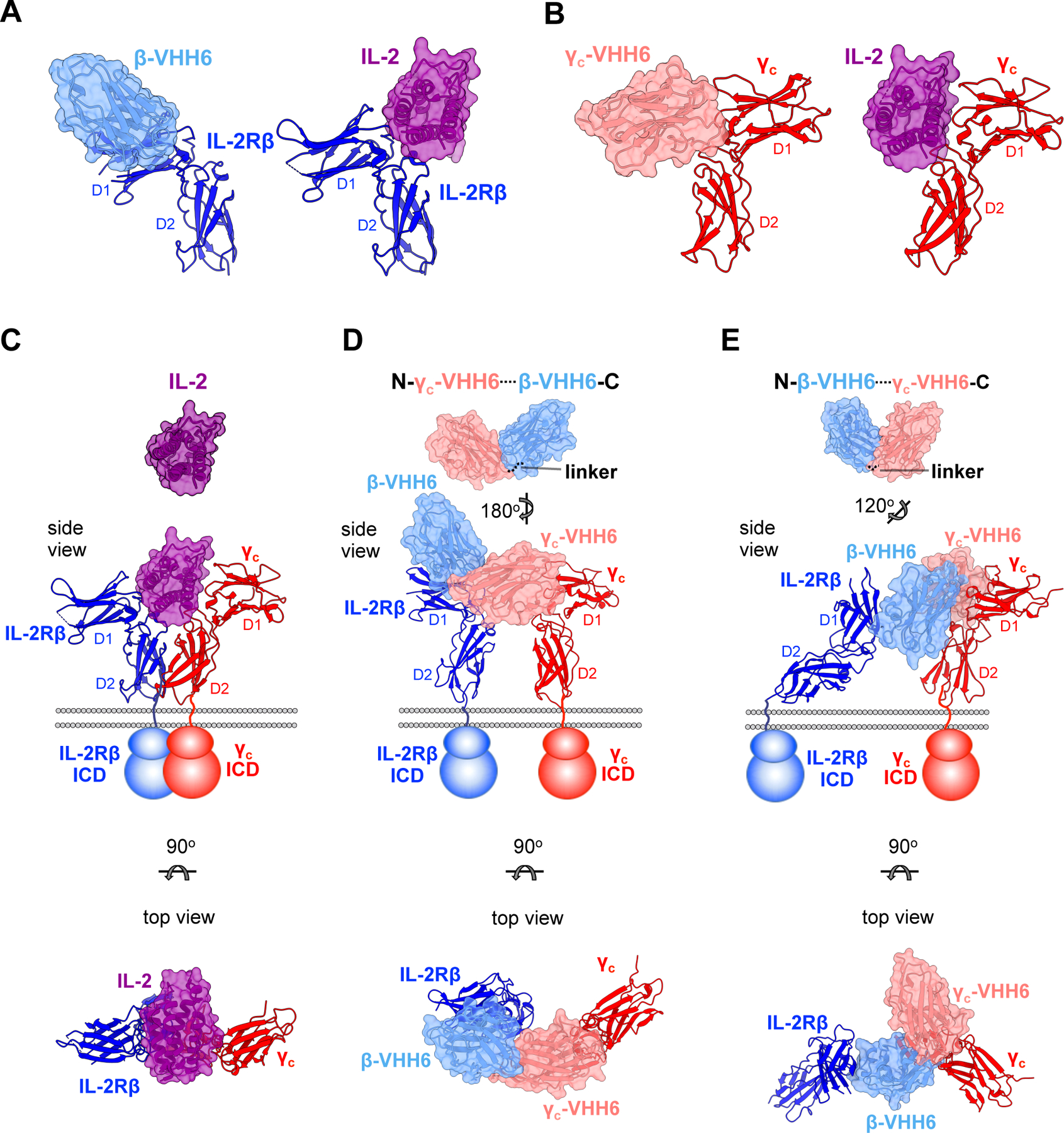
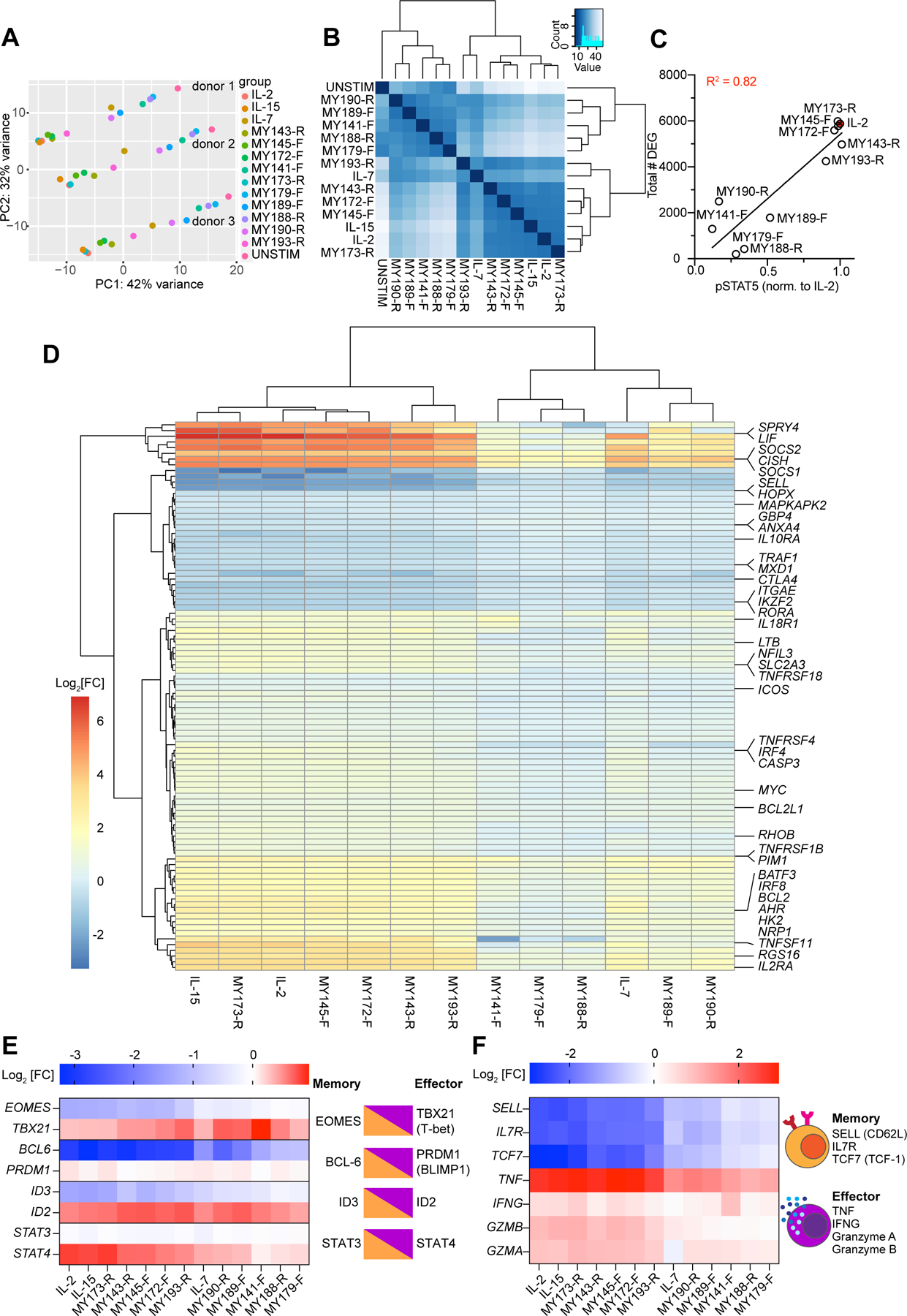
Cytokines are powerful immune modulators that initiate signaling through receptor dimerization, but natural cytokines have structural limitations as therapeutics. We present a strategy to discover cytokine surrogate agonists by using modular ligands that exploit induced proximity and receptor dimer geometry as pharmacological metrics amenable to high-throughput screening. Using VHH and scFv to human interleukin-2/15, type-I interferon, and interleukin-10 receptors, we generated combinatorial matrices of single-chain bispecific ligands that exhibited diverse spectrums of functional activities, including potent inhibition of SARS-CoV-2 by surrogate interferons. Crystal structures of IL-2R:VHH complexes revealed that variation in receptor dimer geometries resulted in functionally diverse signaling outputs. This modular platform enabled engineering of surrogate ligands that compelled assembly of an IL-2R/IL-10R heterodimer, which does not naturally exist, that signaled through pSTAT5 on T and natural killer (NK) cells. This "cytokine med-chem" approach, rooted in principles of induced proximity, is generalizable for discovery of diversified agonists for many ligand-receptor systems.
Keywords: SARS-CoV-2; VHH; bispecific antibodies; cell signaling; cytokine engineering; interferon; interleukin-10; interleukin-2; scFv.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests K.C.G., M.Y., J.R., and Q.L. are co-inventors on a provisional patent 63/306,882 based upon the technology described in this manuscript. K.C.G. is the founder of Synthekine Therapeutics.
Figures
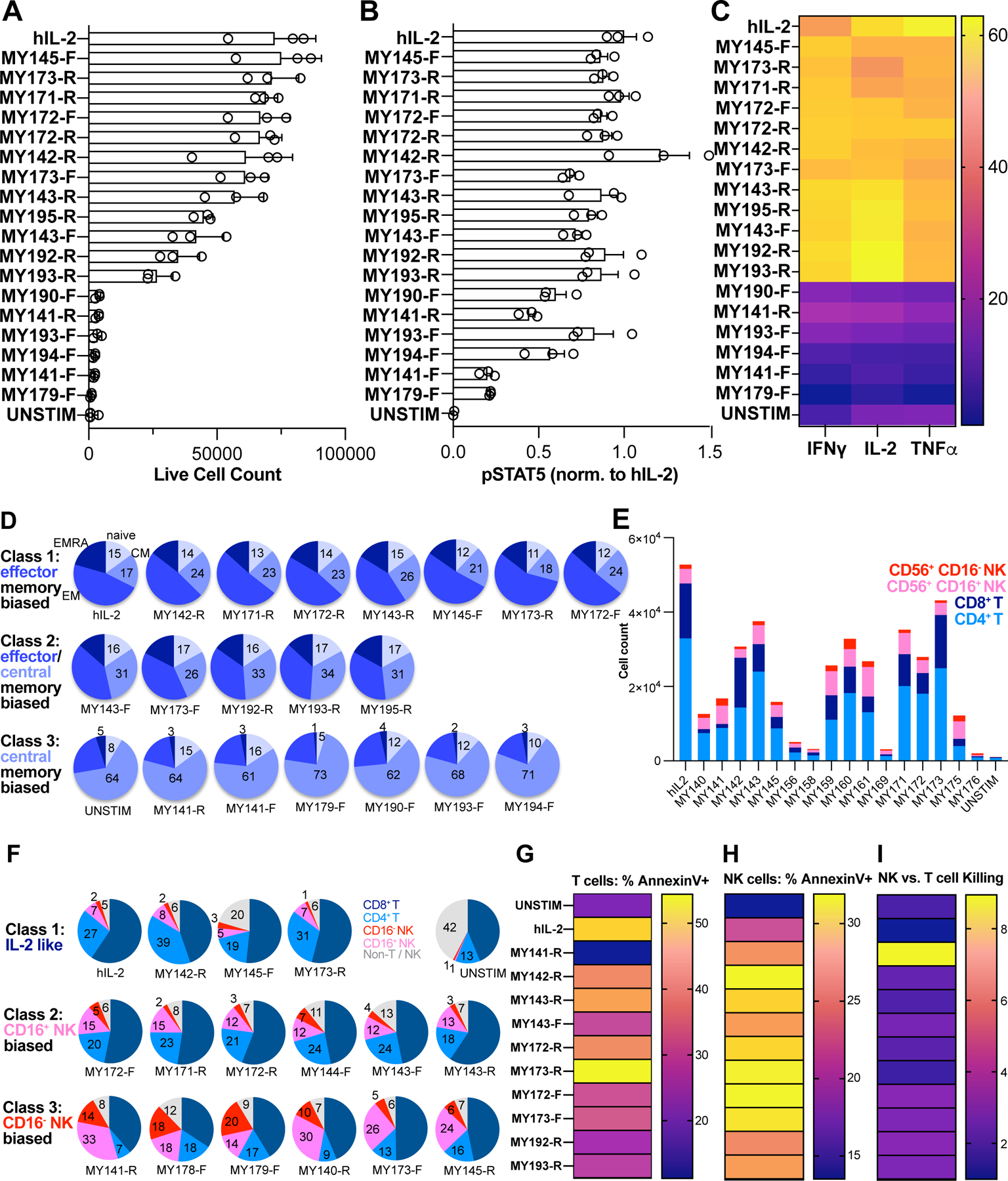
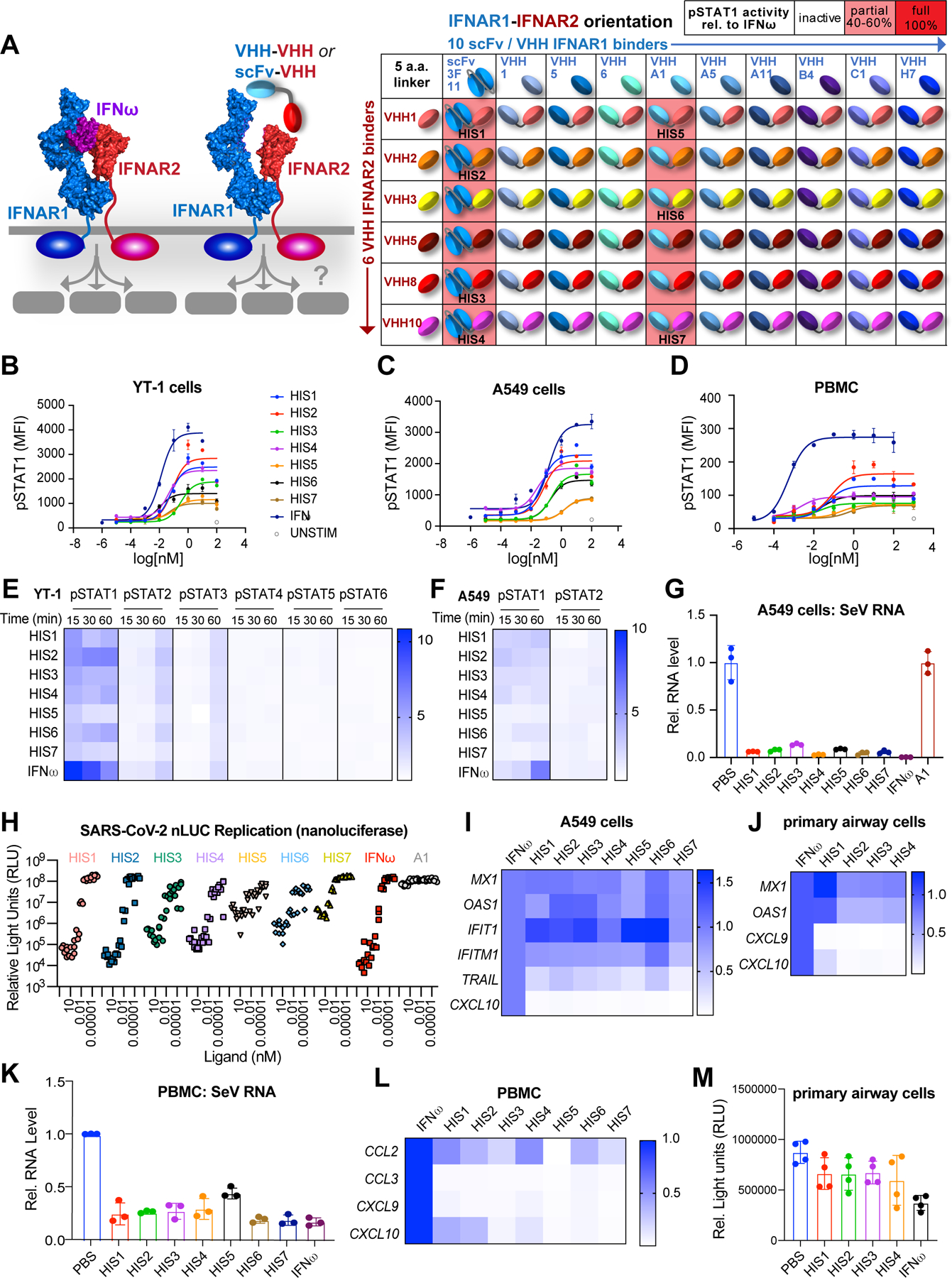
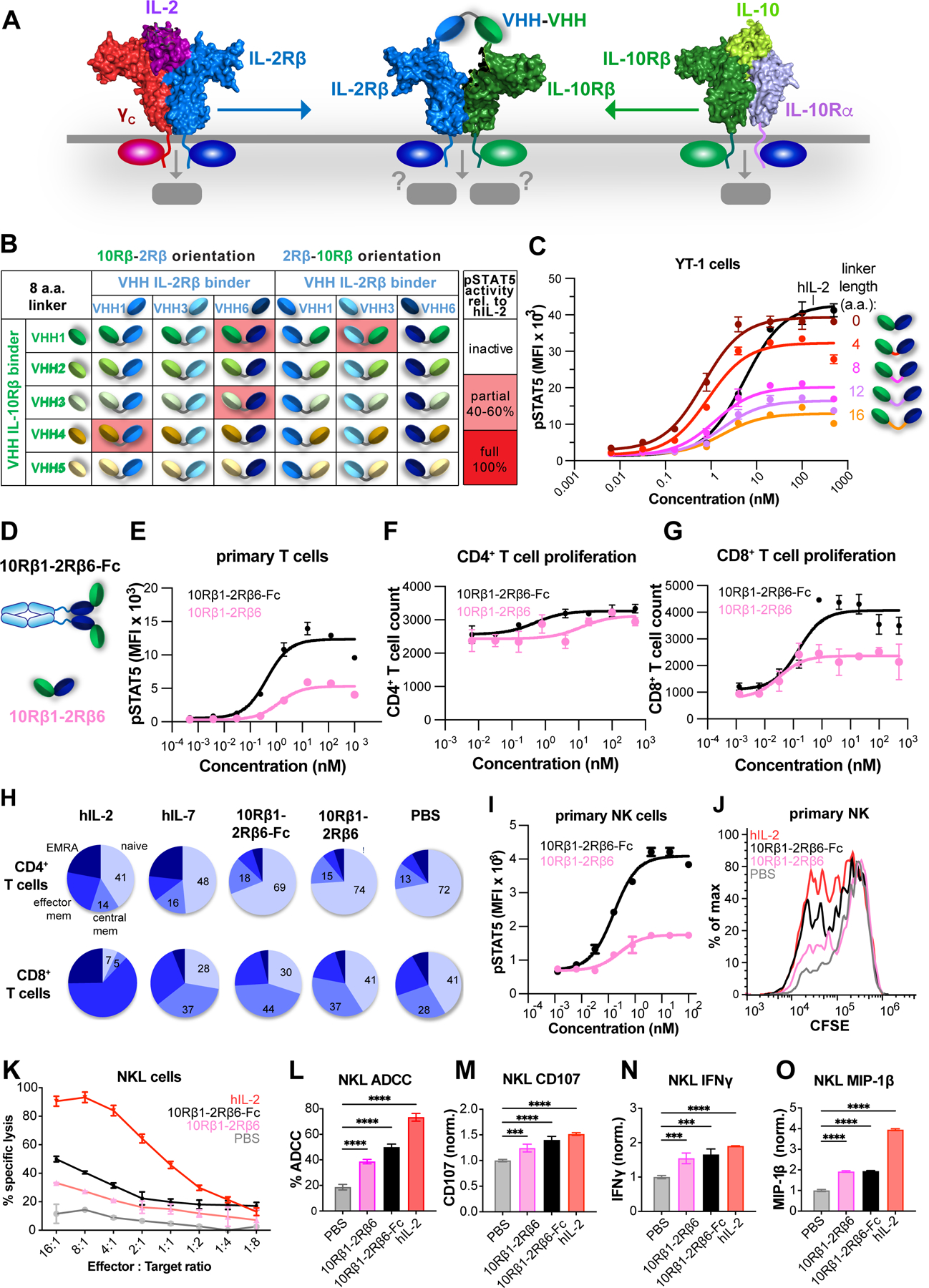
Comment in
-
Surrogate cytokine agonists: promising agents against COVID-19.Signal Transduct Target Ther. 2022 May 6;7(1):150. doi: 10.1038/s41392-022-01015-w. Signal Transduct Target Ther. 2022. PMID: 35523768 Free PMC article. No abstract available.
References
-
- Cardarelli JM, Witte A, and Srinivasan M (2010). Interferon Alpha Receptor 1 Antibodies and Their Uses. US Patent 7662381 B2, filed June 20, 2005, and granted February 16, 2010
-
- Cooper MA, Fehniger TA, and Caligiuri MA (2001). The biology of human natural killercell subsets. Trends Immunol 22, 633–640. - PubMed
-
- Corman SL, and Mohammad RA (2010). Eltrombopag: a novel oral thrombopoietin receptor agonist. Ann Pharmacother 44, 1072–1079. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
