

Transposable Elements: Major Players in Shaping Genomic and Evolutionary Patterns
- PMID: 35326499
- PMCID: PMC8947103
- DOI: 10.3390/cells11061048
Transposable Elements: Major Players in Shaping Genomic and Evolutionary Patterns
Abstract
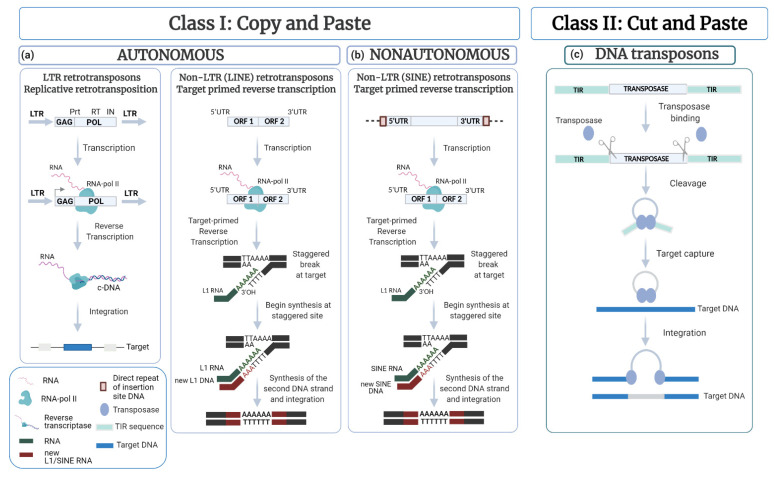
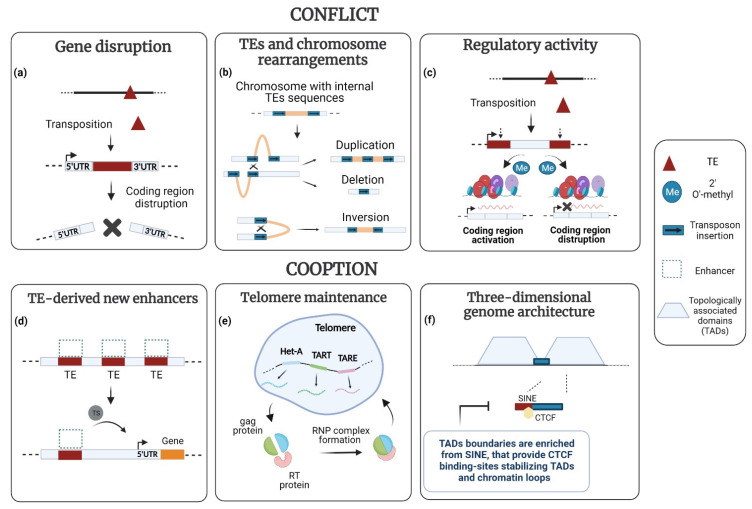
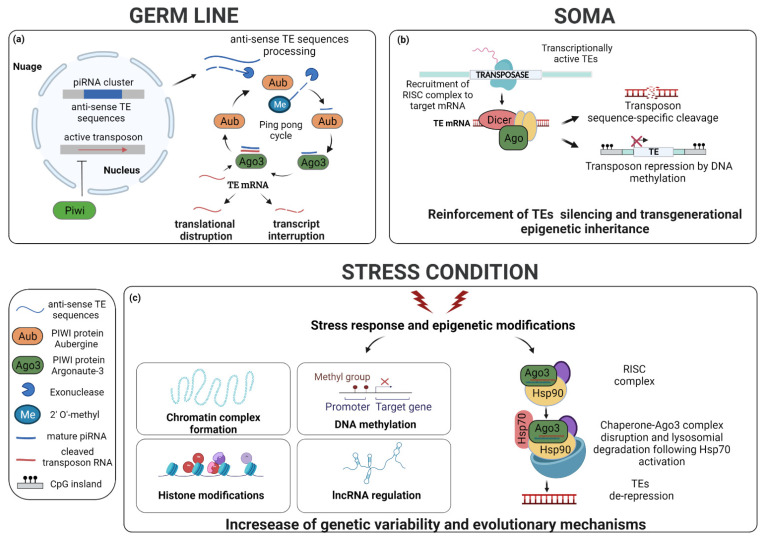
Transposable elements (TEs) are ubiquitous genetic elements, able to jump from one location of the genome to another, in all organisms. For this reason, on the one hand, TEs can induce deleterious mutations, causing dysfunction, disease and even lethality in individuals. On the other hand, TEs can increase genetic variability, making populations better equipped to respond adaptively to environmental change. To counteract the deleterious effects of TEs, organisms have evolved strategies to avoid their activation. However, their mobilization does occur. Usually, TEs are maintained silent through several mechanisms, but they can be reactivated during certain developmental windows. Moreover, TEs can become de-repressed because of drastic changes in the external environment. Here, we describe the 'double life' of TEs, being both 'parasites' and 'symbionts' of the genome. We also argue that the transposition of TEs contributes to two important evolutionary processes: the temporal dynamic of evolution and the induction of genetic variability. Finally, we discuss how the interplay between two TE-dependent phenomena, insertional mutagenesis and epigenetic plasticity, plays a role in the process of evolution.
Keywords: environmental stress; epigenetics; evolution; transposable elements.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Stanley S.M. Speciation, and the fossil record. Prog. Clin. Biol. Res. 1982;96:41–49. - PubMed
-
- Maynard Smith J. Games, Sex, and Evolution. Harvester-Wheatsheaf; Hoboken, NJ, USA: 1988. p. 264.
-
- Gould S.J. Wonderful Life: The Burgess Shale and the Nature of History. W. W. Norton and Company; New York, NY, USA: 1989. p. 347.
-
- Schindewolf O.H. Basic Questions in Paleontology: Geologic Time, Organic Evolution, and Biological Systematics. University of Chicago Press; Chicago, IL, USA: 1993.
-
- Rudwick M.J.S., Cuvier G. Fossil Bones, and Geological Catastrophes. University of Chicago Press; Chicago, IL, USA: 1997. p. 301.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
