

New Glycosylated Polyene Macrolides: Refining the Ore from Genome Mining
- PMID: 35326797
- PMCID: PMC8944477
- DOI: 10.3390/antibiotics11030334
New Glycosylated Polyene Macrolides: Refining the Ore from Genome Mining
Abstract
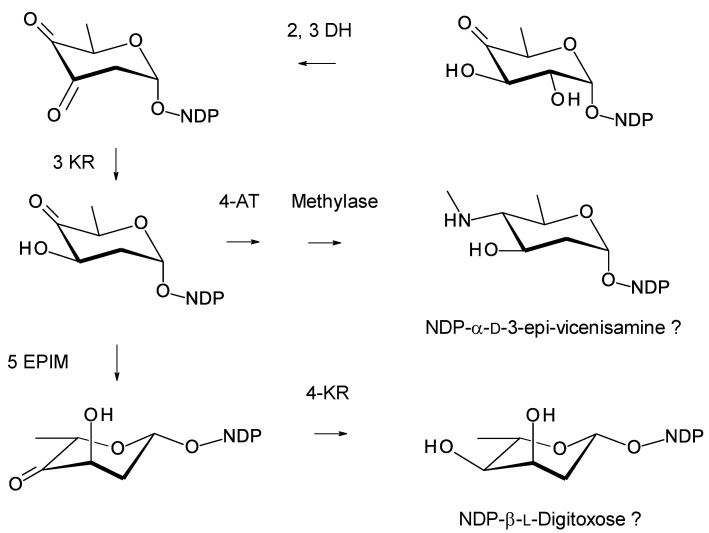
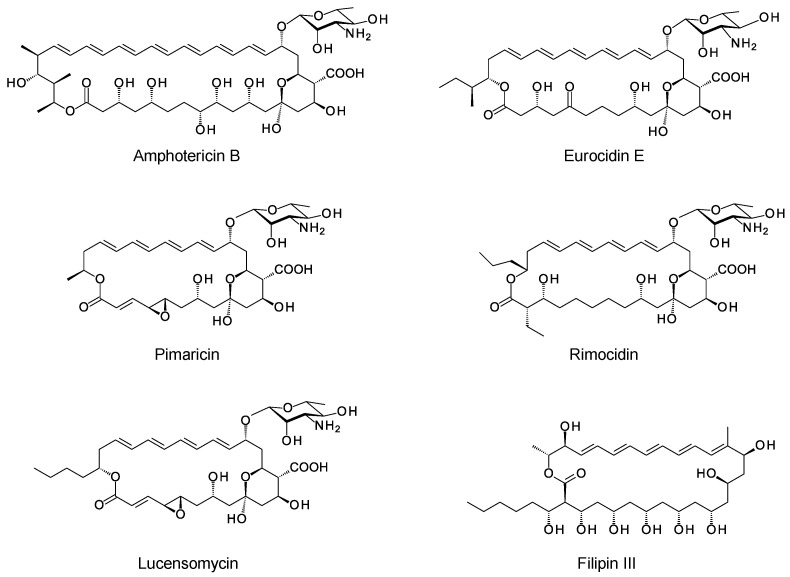
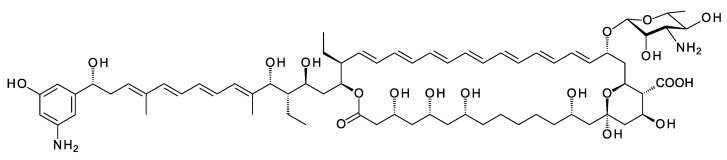
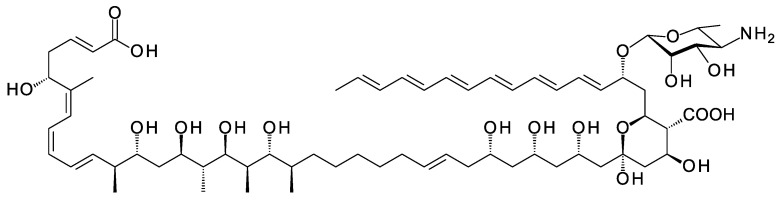
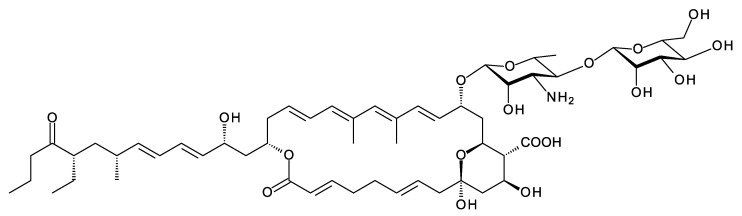
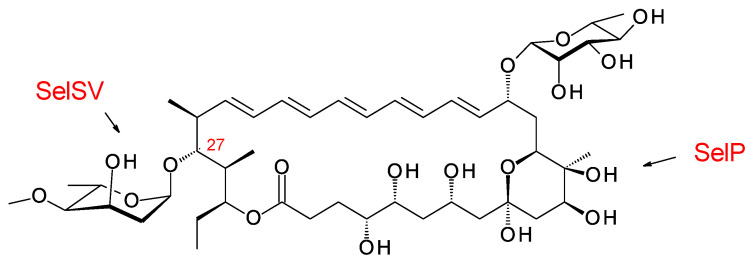
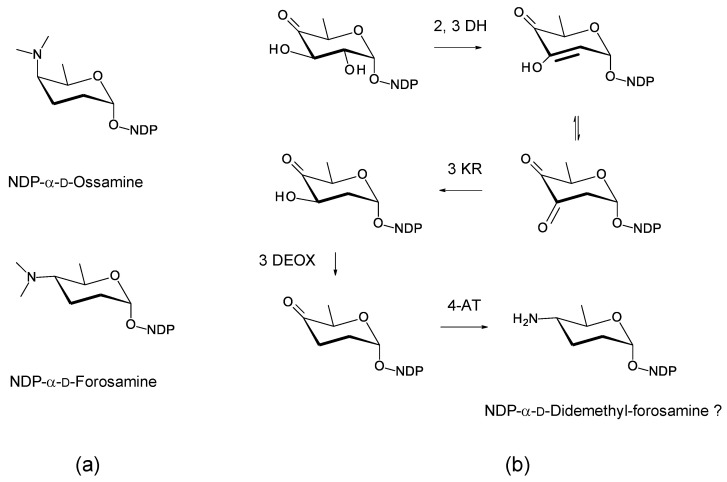
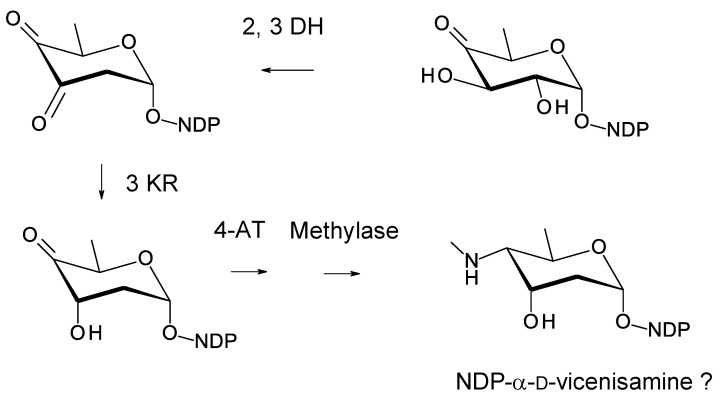
Glycosylated polyene macrolides include effective antifungal agents, such as pimaricin, nystatin, candicidin, and amphotericin B. For the treatment of systemic mycoses, amphotericin B has been described as a gold-standard antibiotic because of its potent activity against a broad spectrum of fungal pathogens, which do not readily become resistant. However, amphotericin B has severe toxic side effects, and the development of safer alternatives remains an important objective. One approach towards obtaining such compounds is to discover new related natural products. Advances in next-generation sequencing have delivered a wealth of microbial genome sequences containing polyene biosynthetic gene clusters. These typically encode a modular polyketide synthase that catalyzes the assembly of the aglycone core, a cytochrome P450 that oxidizes a methyl branch to a carboxyl group, and additional enzymes for synthesis and attachment of a single mycosamine sugar residue. In some cases, further P450s catalyze epoxide formation or hydroxylation within the macrolactone. Bioinformatic analyses have identified over 250 of these clusters. Some are predicted to encode potentially valuable new polyenes that have not been uncovered by traditional screening methods. Recent experimental studies have characterized polyenes with new polyketide backbones, previously unknown late oxygenations, and additional sugar residues that increase water-solubility and reduce hemolytic activity. Here we review these studies and assess how this new knowledge can help to prioritize silent polyene clusters for further investigation. This approach should improve the chances of discovering better antifungal antibiotics.
Keywords: antifungal antibiotics; biosynthetic gene clusters; genome mining; glycosylated polyene macrolide.
Conflict of interest statement
The authors declare no conflict of interest.
Figures























References
-
- Omura S., Tanaka H. Production, Structure and Antifungal Activity of Polyene Macrolides. In: Omura S., editor. Macrolide Antibiotics, Chemistry, Biology and Practice. Academic Press; New York, NY, USA: 1986. pp. 351–404.
-
- Abu-Salah K.M. Amphotericin B: An update. Br. J. Biomed. Sci. 1996;53:8757689. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
