

The Role of Membrane Lipids in Light-Activation of Drosophila TRP Channels
- PMID: 35327573
- PMCID: PMC8945425
- DOI: 10.3390/biom12030382
The Role of Membrane Lipids in Light-Activation of Drosophila TRP Channels
Abstract
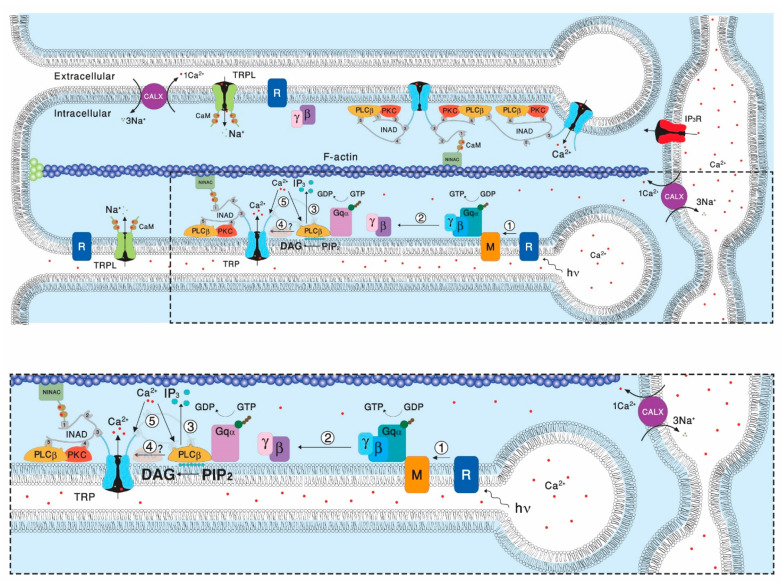
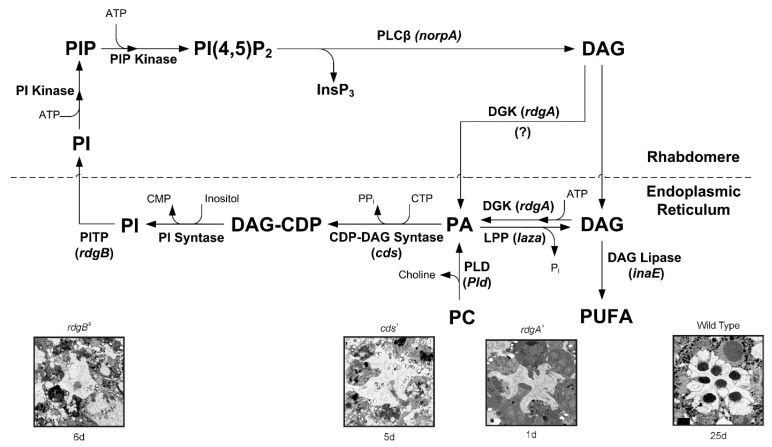
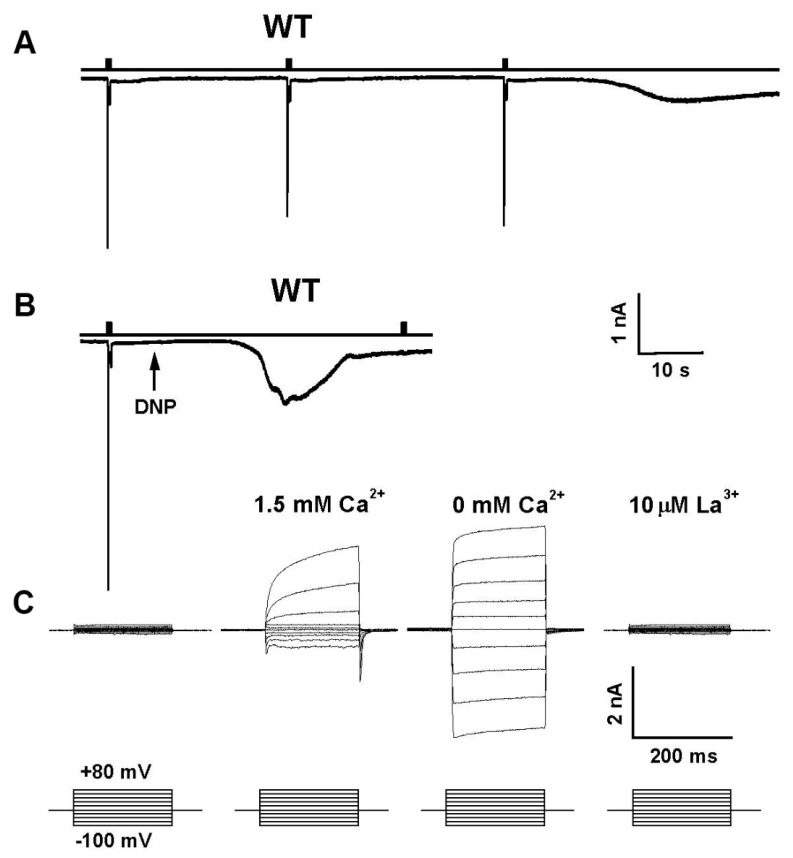
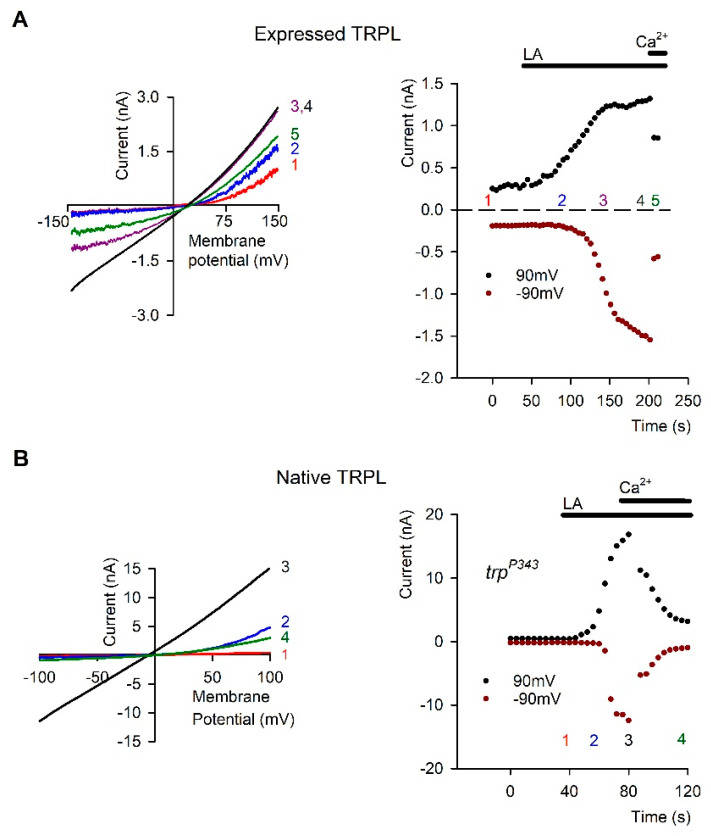
Transient Receptor Potential (TRP) channels constitute a large superfamily of polymodal channel proteins with diverse roles in many physiological and sensory systems that function both as ionotropic and metabotropic receptors. From the early days of TRP channel discovery, membrane lipids were suggested to play a fundamental role in channel activation and regulation. A prominent example is the Drosophila TRP and TRP-like (TRPL) channels, which are predominantly expressed in the visual system of Drosophila. Light activation of the TRP and TRPL channels, the founding members of the TRP channel superfamily, requires activation of phospholipase Cβ (PLC), which hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) into Diacylglycerol (DAG) and Inositol 1, 4,5-trisphosphate (IP3). However, the events required for channel gating downstream of PLC activation are still under debate and led to several hypotheses regarding the mechanisms by which lipids gate the channels. Despite many efforts, compelling evidence of the involvement of DAG accumulation, PIP2 depletion or IP3-mediated Ca2+ release in light activation of the TRP/TRPL channels are still lacking. Exogeneous application of poly unsaturated fatty acids (PUFAs), a product of DAG hydrolysis was demonstrated as an efficient way to activate the Drosophila TRP/TRPL channels. However, compelling evidence for the involvement of PUFAs in physiological light-activation of the TRP/TRPL channels is still lacking. Light-induced mechanical force generation was measured in photoreceptor cells prior to channel opening. This mechanical force depends on PLC activity, suggesting that the enzymatic activity of PLC converting PIP2 into DAG generates membrane tension, leading to mechanical gating of the channels. In this review, we will present the roles of membrane lipids in light activation of Drosophila TRP channels and present the many advantages of this model system in the exploration of TRP channel activation under physiological conditions.
Keywords: Diacylglycerol (DAG); Diacylglycerol kinase (DGK); Drosophila TRP channel; TRPL channel; cholesterol; ergosterol; methyl-β-cyclodextrin; phospholipase Cβ; poly unsaturated fatty acids (PUFAs).
Conflict of interest statement
Authors declare no conflict of interest.
Figures
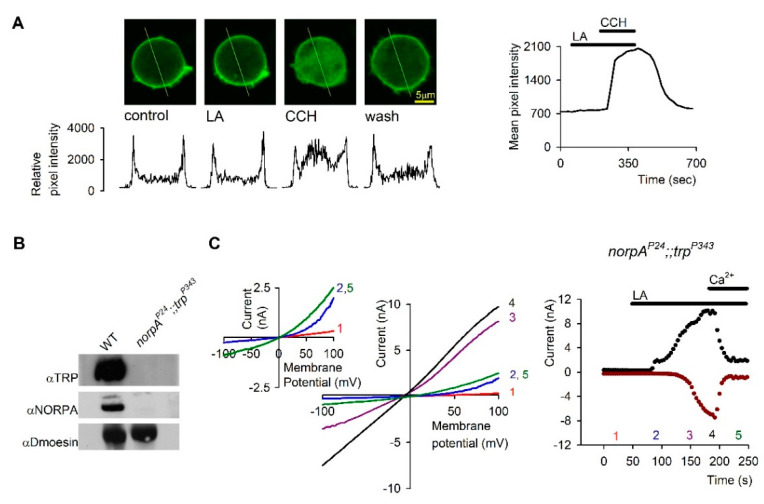
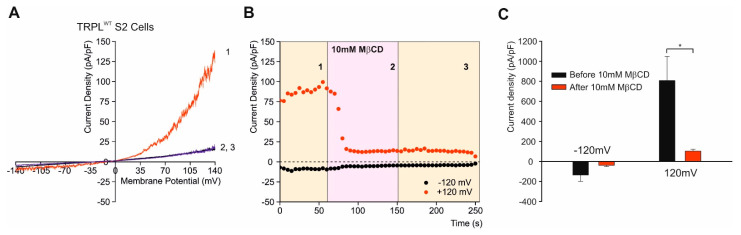
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
