

T-cell Receptor Is a Threshold Detector: Sub- and Supra-Threshold Stochastic Resonance in TCR-MHC Clusters on the Cell Surface
- PMID: 35327900
- PMCID: PMC8946872
- DOI: 10.3390/e24030389
T-cell Receptor Is a Threshold Detector: Sub- and Supra-Threshold Stochastic Resonance in TCR-MHC Clusters on the Cell Surface
Abstract
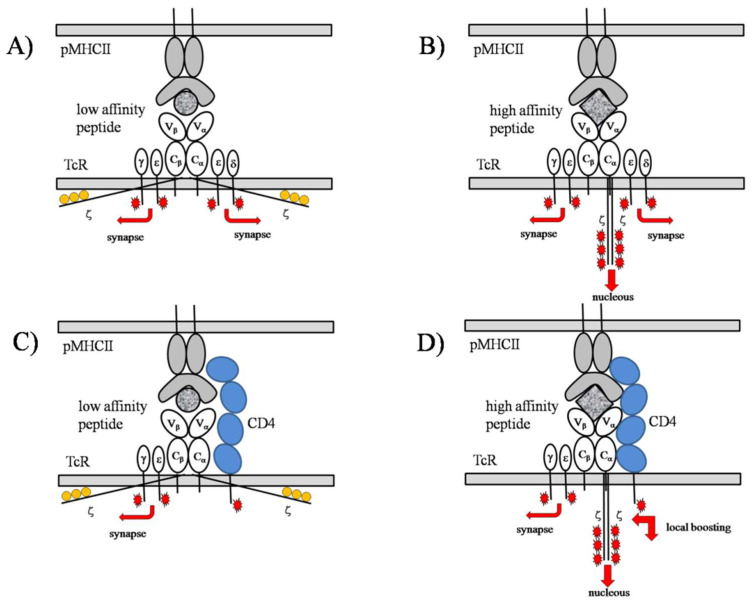
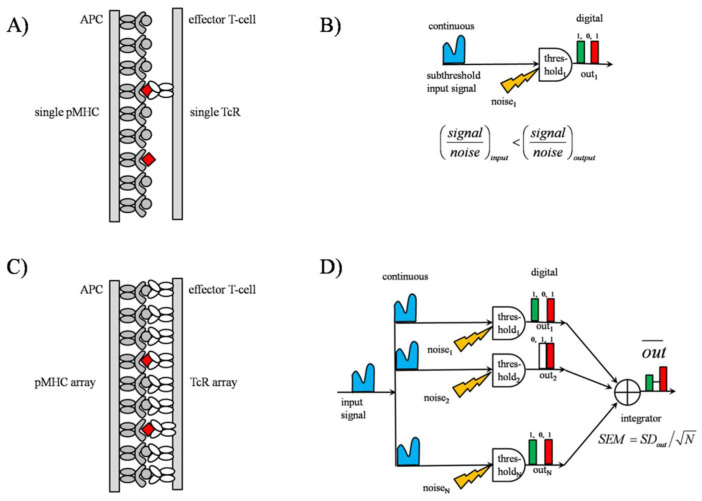
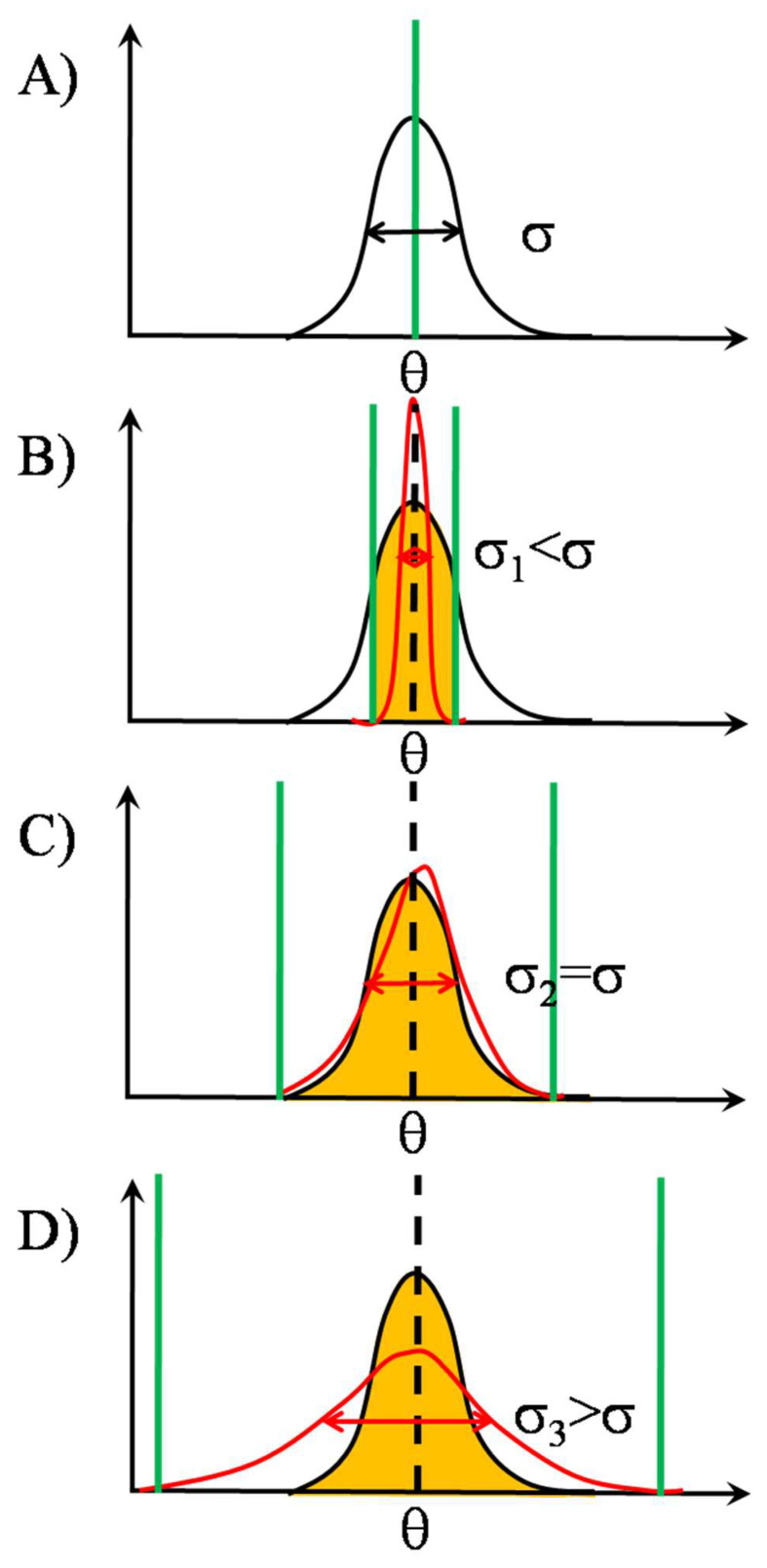
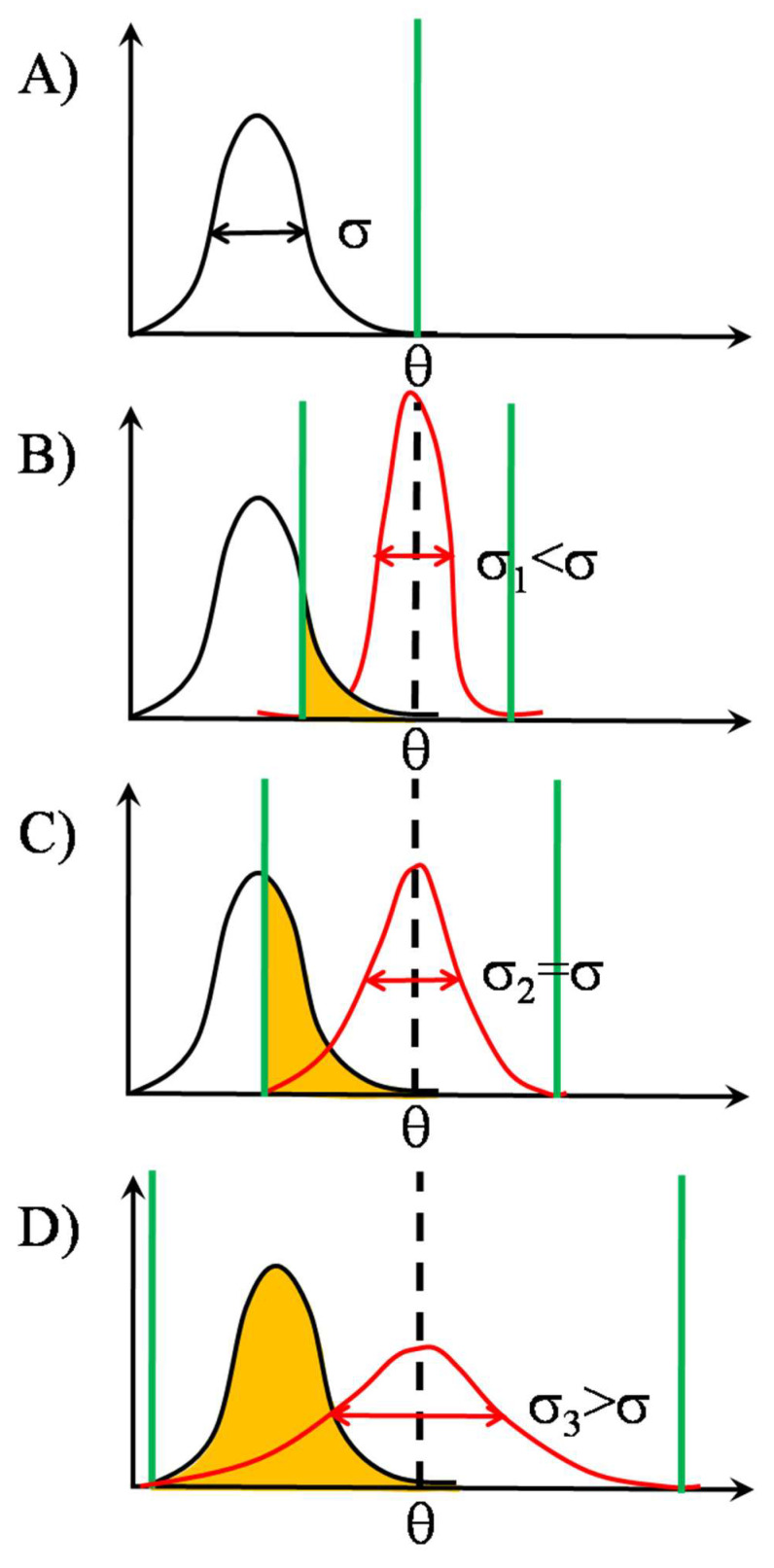
Stochastic resonance in clusters of major histocompatibility molecules is extended by a more detailed description of adaptive thresholding and by applying the notion of suprathreshold stochastic resonance as a stochastically quantizing encoder of transmembrane signaling downstream of major histocompatibility molecules and T-cell receptors on the side of presenting and recognizing cells, respectively. The adaptive nature of thresholding is partly explained by a mirroring of the noncognate-cognate dichotomy shown by the T-cell receptor structure and the kinetic-segregation model of the onset of T-cell receptor triggering. Membrane clusters of major histocompatibility molecules and T-cell receptors on their host cells are envisioned as places of the temporal encoding of downstream signals via the suprathreshold stochastic resonance process. The ways of optimization of molecular prostheses, such as chimeric antigen receptors against cancer in transmembrane signaling, are suggested in the framework of suprathreshold stochastic resonance. The analogy between Förster resonance energy transfer and suprathreshold stochastic resonance for information transfer is also discussed. The overlap integral for energy transfer parallels the mutual information transferred by suprathreshold stochastic resonance.
Keywords: CD8/CD4; Förster resonance energy transfer (FRET); MHCI/MHCII; TCR; chimeric antigen receptor (CAR) optimization; correlation; distributed signal detection; mutual information; noncognate–cognate discrimination; pattern recognition.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
Adaptive threshold-stochastic resonance (AT-SR) in MHC clusters on the cell surface.Immunol Lett. 2020 Jan;217:65-71. doi: 10.1016/j.imlet.2019.11.006. Epub 2019 Nov 15. Immunol Lett. 2020. PMID: 31738956 Review.
-
Functional evidence for TCR-intrinsic specificity for MHCII.Proc Natl Acad Sci U S A. 2016 Mar 15;113(11):3000-5. doi: 10.1073/pnas.1518499113. Epub 2016 Feb 1. Proc Natl Acad Sci U S A. 2016. PMID: 26831112 Free PMC article.
-
CD4 and CD8 accessory molecules function through interactions with major histocompatibility complex molecules which are not directly associated with the T cell receptor-antigen complex.Eur J Immunol. 1991 Oct;21(10):2507-15. doi: 10.1002/eji.1830211030. Eur J Immunol. 1991. PMID: 1915555
-
Thymocyte development in major histocompatibility complex-deficient mice: evidence for stochastic commitment to the CD4 and CD8 lineages.Proc Natl Acad Sci U S A. 1993 Nov 15;90(22):10739-43. doi: 10.1073/pnas.90.22.10739. Proc Natl Acad Sci U S A. 1993. PMID: 7902569 Free PMC article.
-
Interactions between MHC molecules and co-receptors of the TCR.Curr Opin Immunol. 2002 Feb;14(1):75-83. doi: 10.1016/s0952-7915(01)00300-4. Curr Opin Immunol. 2002. PMID: 11790535 Review.
Cited by
-
Robust neural networks using stochastic resonance neurons.Commun Eng. 2024 Nov 13;3(1):169. doi: 10.1038/s44172-024-00314-0. Commun Eng. 2024. PMID: 39537964 Free PMC article.
References
-
- Damjanovich S., Bene L., Matkó J., Mátyus L., Krasznai Z., Szabó G., Jr., Pieri C., Gáspár R., Jr., Szöllősi J. Two-dimensional receptor patterns in the plasma membrane of cells. A critical evaluation of their identification, origin and information content. Biophys. Chem. 1999;82:99–108. doi: 10.1016/S0301-4622(99)00109-X. - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
