

Epitranscriptomic Reprogramming Is Required to Prevent Stress and Damage from Acetaminophen
- PMID: 35327975
- PMCID: PMC8955276
- DOI: 10.3390/genes13030421
Epitranscriptomic Reprogramming Is Required to Prevent Stress and Damage from Acetaminophen
Abstract
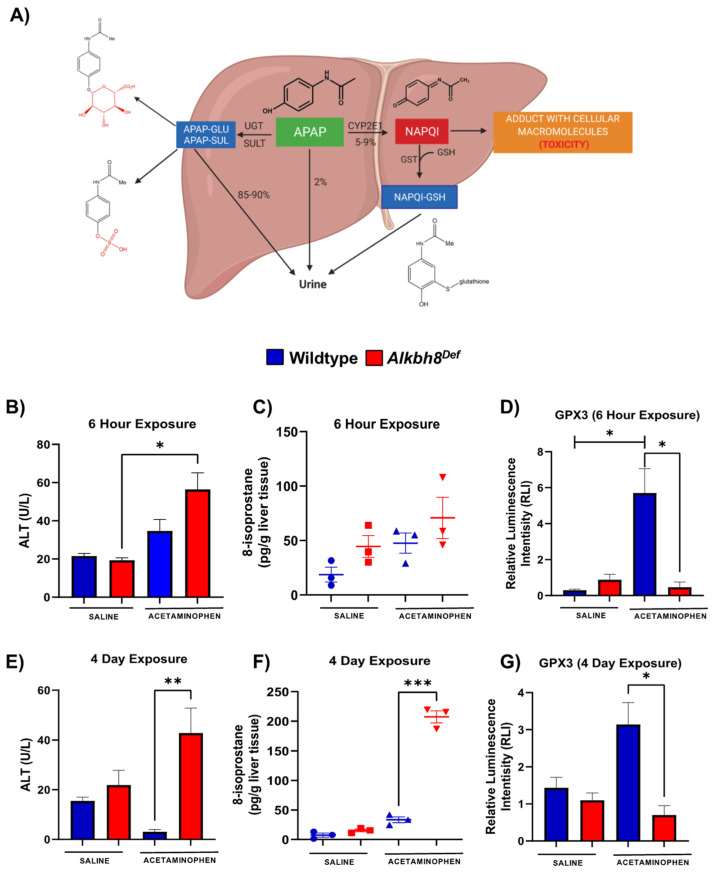
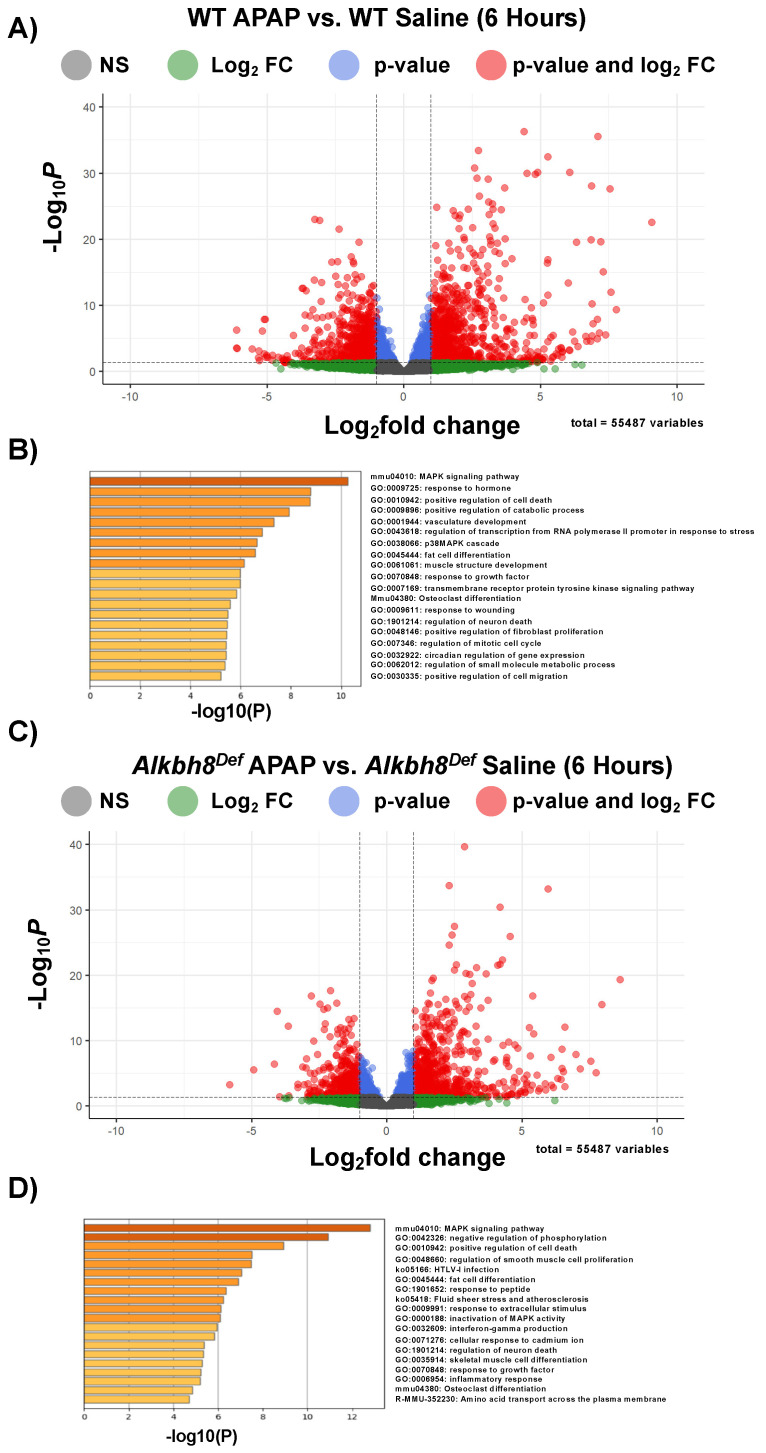
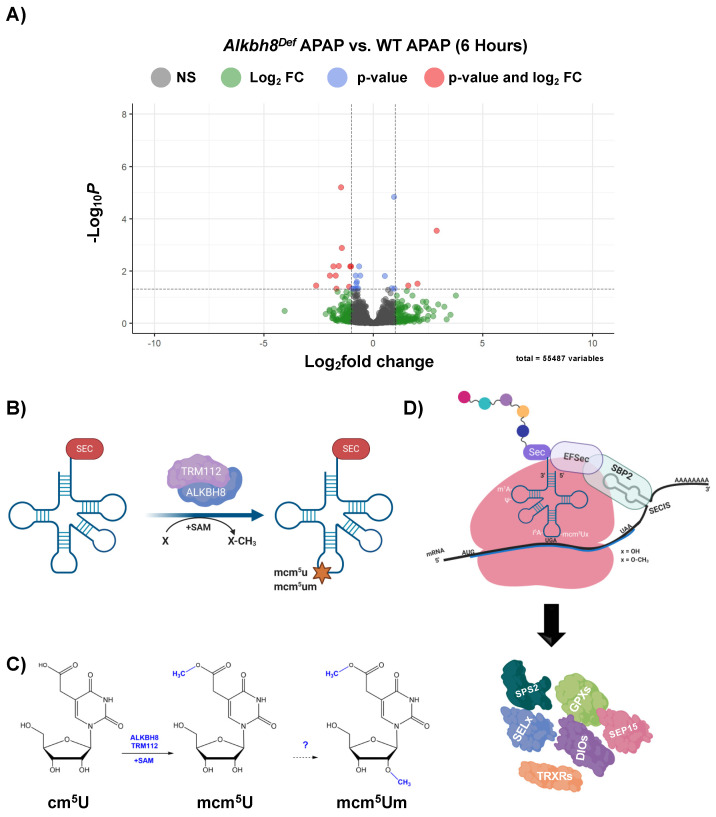
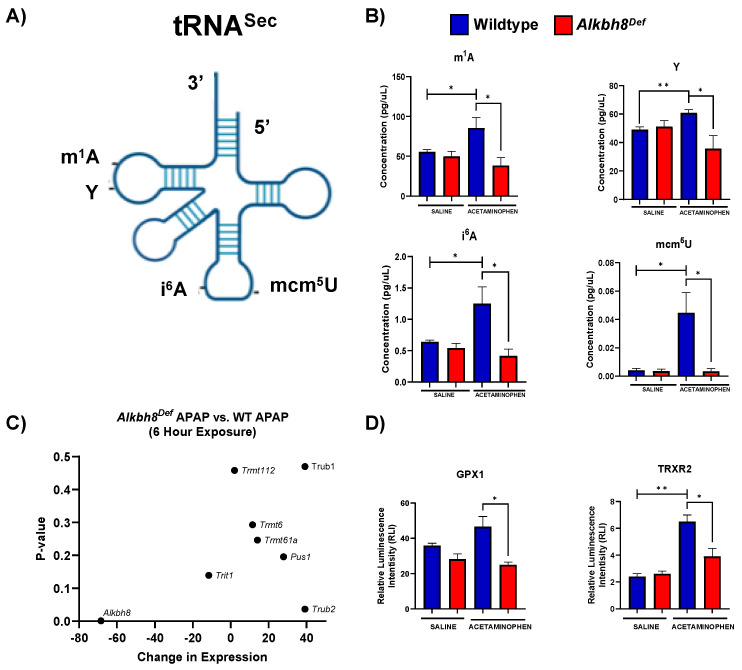
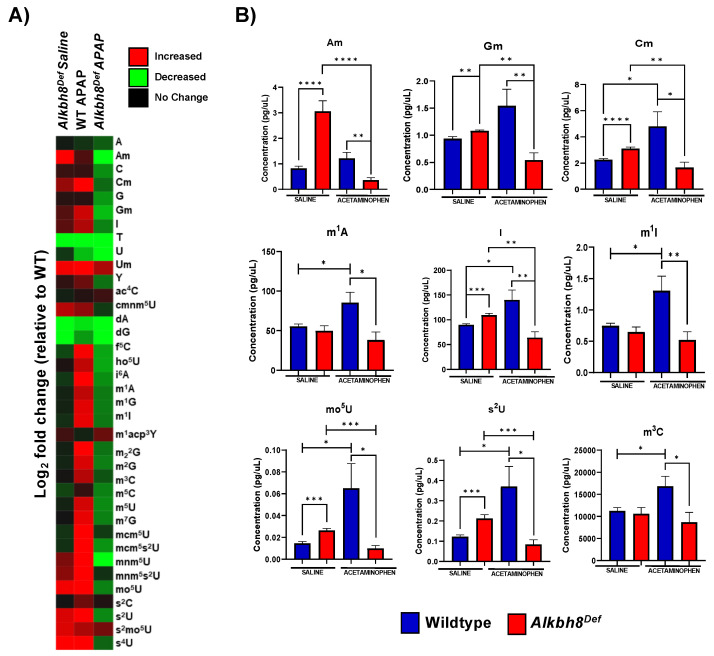
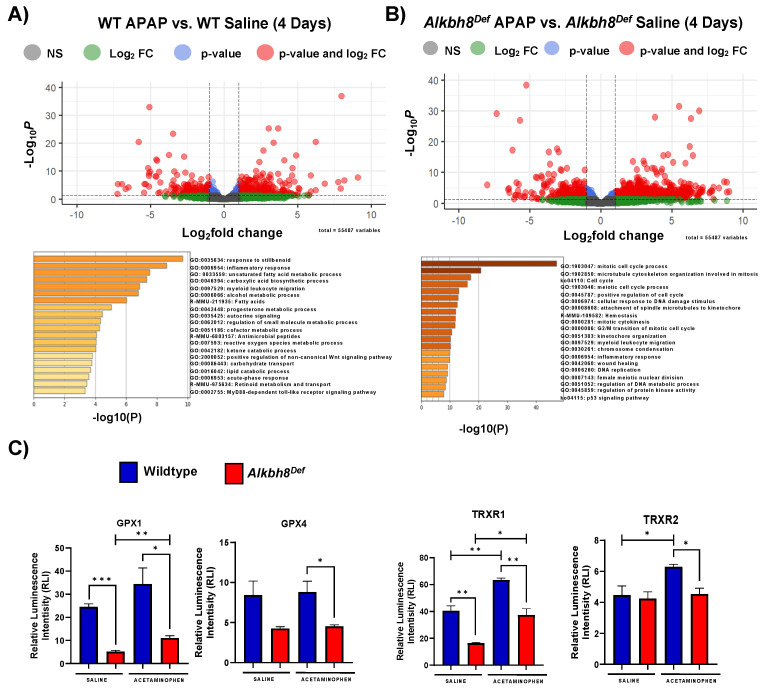
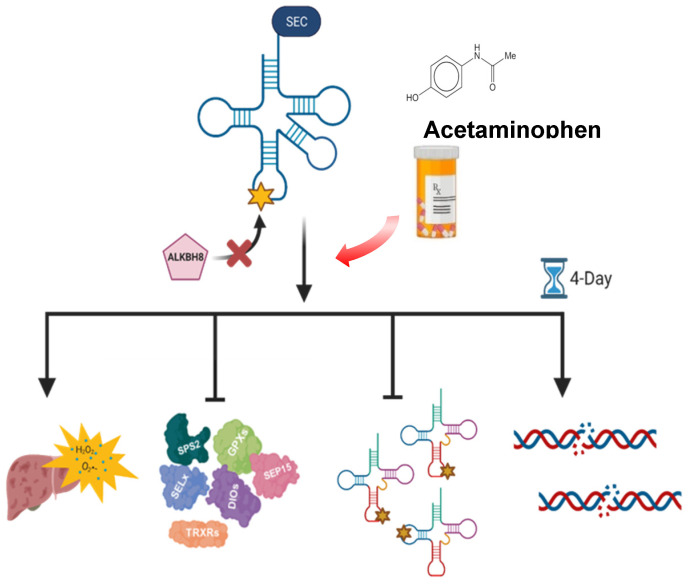
Epitranscriptomic marks, in the form of enzyme catalyzed RNA modifications, play important gene regulatory roles in response to environmental and physiological conditions. However, little is known with respect to how acute toxic doses of pharmaceuticals influence the epitranscriptome. Here we define how acetaminophen (APAP) induces epitranscriptomic reprogramming and how the writer Alkylation Repair Homolog 8 (Alkbh8) plays a key gene regulatory role in the response. Alkbh8 modifies tRNA selenocysteine (tRNASec) to translationally regulate the production of glutathione peroxidases (Gpx's) and other selenoproteins, with Gpx enzymes known to play protective roles during APAP toxicity. We demonstrate that APAP increases toxicity and markers of damage, and decreases selenoprotein levels in Alkbh8 deficient mouse livers, when compared to wildtype. APAP also promotes large scale reprogramming of many RNA marks comprising the liver tRNA epitranscriptome including: 5-methoxycarbonylmethyluridine (mcm5U), isopentenyladenosine (i6A), pseudouridine (Ψ), and 1-methyladenosine (m1A) modifications linked to tRNASec and many other tRNA's. Alkbh8 deficiency also leads to wide-spread epitranscriptomic dysregulation in response to APAP, demonstrating that a single writer defect can promote downstream changes to a large spectrum of RNA modifications. Our study highlights the importance of RNA modifications and translational responses to APAP, identifies writers as key modulators of stress responses in vivo and supports the idea that the epitranscriptome may play important roles in responses to pharmaceuticals.
Keywords: Alkbh8; RNA modification; acetaminophen; epitranscriptomic; stress response; tRNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Selenoproteins and the senescence-associated epitranscriptome.Exp Biol Med (Maywood). 2022 Dec;247(23):2090-2102. doi: 10.1177/15353702221116592. Epub 2022 Aug 29. Exp Biol Med (Maywood). 2022. PMID: 36036467 Free PMC article. Review.
-
The epitranscriptomic writer ALKBH8 drives tolerance and protects mouse lungs from the environmental pollutant naphthalene.Epigenetics. 2020 Oct;15(10):1121-1138. doi: 10.1080/15592294.2020.1750213. Epub 2020 Apr 17. Epigenetics. 2020. PMID: 32303148 Free PMC article.
-
Loss of epitranscriptomic control of selenocysteine utilization engages senescence and mitochondrial reprogramming☆.Redox Biol. 2020 Jan;28:101375. doi: 10.1016/j.redox.2019.101375. Epub 2019 Nov 11. Redox Biol. 2020. PMID: 31765888 Free PMC article.
-
Alkbh8 Regulates Selenocysteine-Protein Expression to Protect against Reactive Oxygen Species Damage.PLoS One. 2015 Jul 6;10(7):e0131335. doi: 10.1371/journal.pone.0131335. eCollection 2015. PLoS One. 2015. PMID: 26147969 Free PMC article.
-
Epitranscriptomic systems regulate the translation of reactive oxygen species detoxifying and disease linked selenoproteins.Free Radic Biol Med. 2019 Nov 1;143:573-593. doi: 10.1016/j.freeradbiomed.2019.08.030. Epub 2019 Aug 30. Free Radic Biol Med. 2019. PMID: 31476365 Free PMC article. Review.
Cited by
-
An Updated Review on the Metabolite (AM404)-Mediated Central Mechanism of Action of Paracetamol (Acetaminophen): Experimental Evidence and Potential Clinical Impact.J Pain Res. 2023 Mar 29;16:1081-1094. doi: 10.2147/JPR.S393809. eCollection 2023. J Pain Res. 2023. PMID: 37016715 Free PMC article. Review.
-
Disruption of the mouse liver epitranscriptome by long-term aroclor 1260 exposure.Environ Toxicol Pharmacol. 2023 Jun;100:104138. doi: 10.1016/j.etap.2023.104138. Epub 2023 May 1. Environ Toxicol Pharmacol. 2023. PMID: 37137421 Free PMC article.
-
The biological function of demethylase ALKBH1 and its role in human diseases.Heliyon. 2024 Jun 24;10(13):e33489. doi: 10.1016/j.heliyon.2024.e33489. eCollection 2024 Jul 15. Heliyon. 2024. PMID: 39040364 Free PMC article. Review.
-
Selenoproteins and the senescence-associated epitranscriptome.Exp Biol Med (Maywood). 2022 Dec;247(23):2090-2102. doi: 10.1177/15353702221116592. Epub 2022 Aug 29. Exp Biol Med (Maywood). 2022. PMID: 36036467 Free PMC article. Review.
-
Recharacterization of RSL3 reveals that the selenoproteome is a druggable target in colorectal cancer.bioRxiv [Preprint]. 2024 Aug 27:2024.03.29.587381. doi: 10.1101/2024.03.29.587381. bioRxiv. 2024. Update in: Cancer Res. 2025 Aug 1;85(15):2788-2804. doi: 10.1158/0008-5472.CAN-24-3478. PMID: 38617233 Free PMC article. Updated. Preprint.
References
-
- Cai W.M., Chionh Y.H., Hia F., Gu C., Kellner S., McBee M.E., Ng C.S., Ling Y., Pang J., Prestwich E.G., et al. A platform for discovery and quantification of modified ribonucleosides in RNA: Application to stress-induced reprogramming of tRNA modificiations. Methods Enzymol. 2016;560:29–71. - PMC - PubMed
-
- Begley U., Sosa M.S., Avivar-Valderas A., Patil A., Endres L., Estrada Y., Chan C.T., Su D., Dedon P.C., Aguirre-Ghiso J.A., et al. A human tRNA methyltransferase 9-like protein prevents tumour growth by regulating LIN9 and HIF1-α. EMBO Mol. Med. 2013;5:366–383. doi: 10.1002/emmm.201201161. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
