

The Rationale for the Dual-Targeting Therapy for RSK2 and AKT in Multiple Myeloma
- PMID: 35328342
- PMCID: PMC8949999
- DOI: 10.3390/ijms23062919
The Rationale for the Dual-Targeting Therapy for RSK2 and AKT in Multiple Myeloma
Abstract
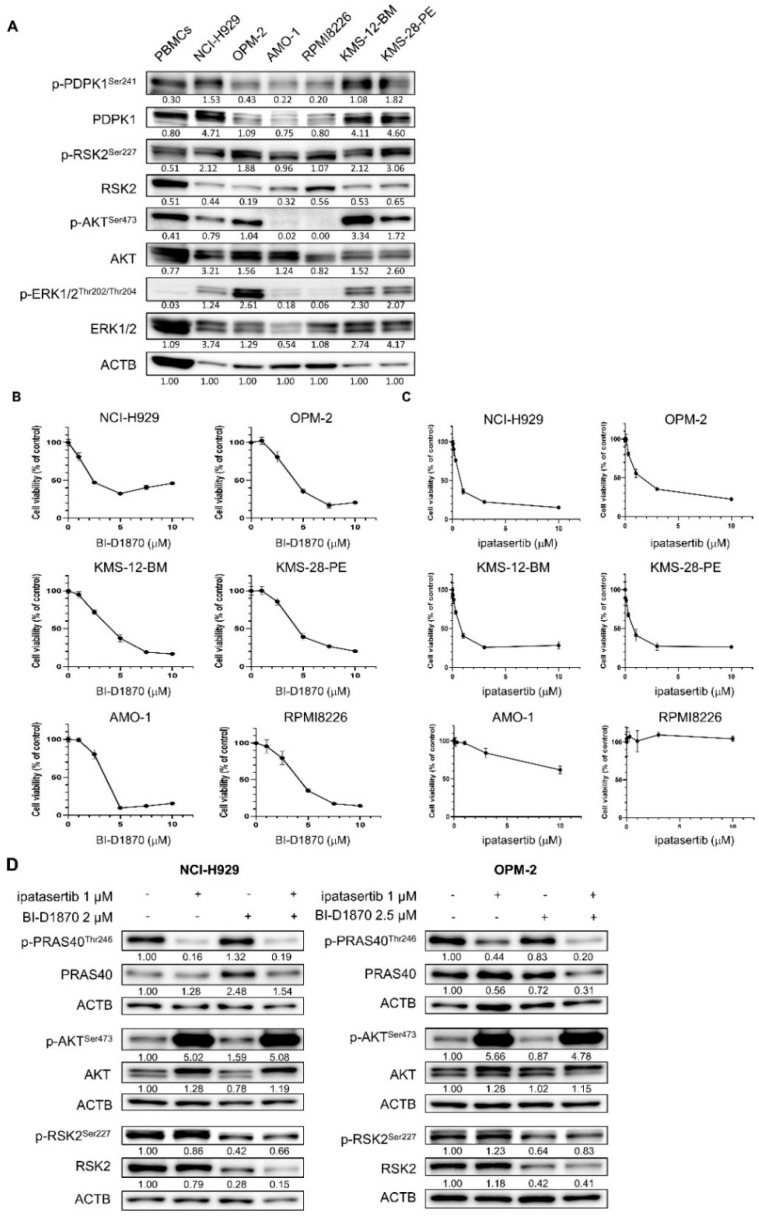
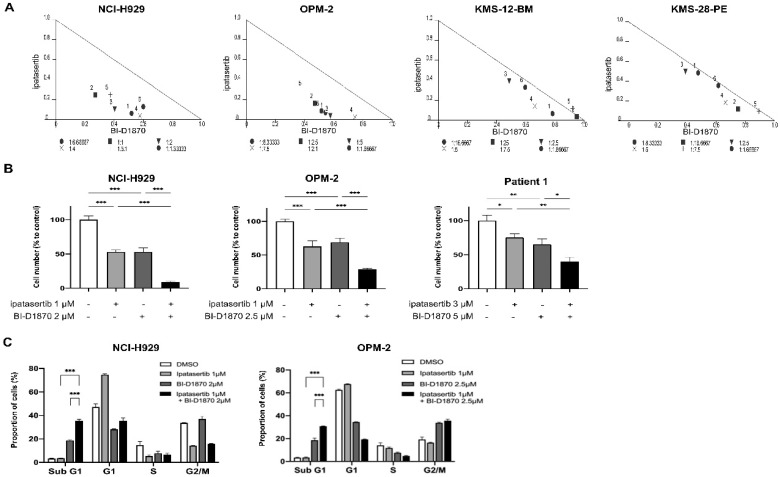
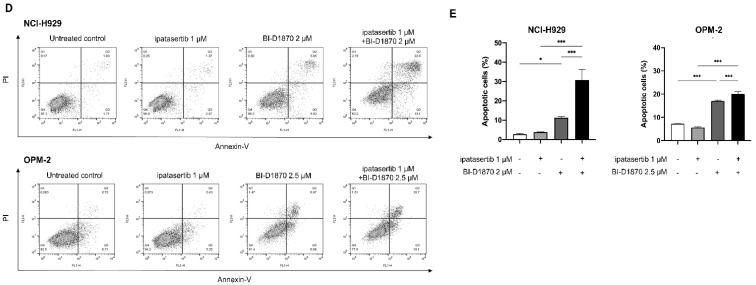
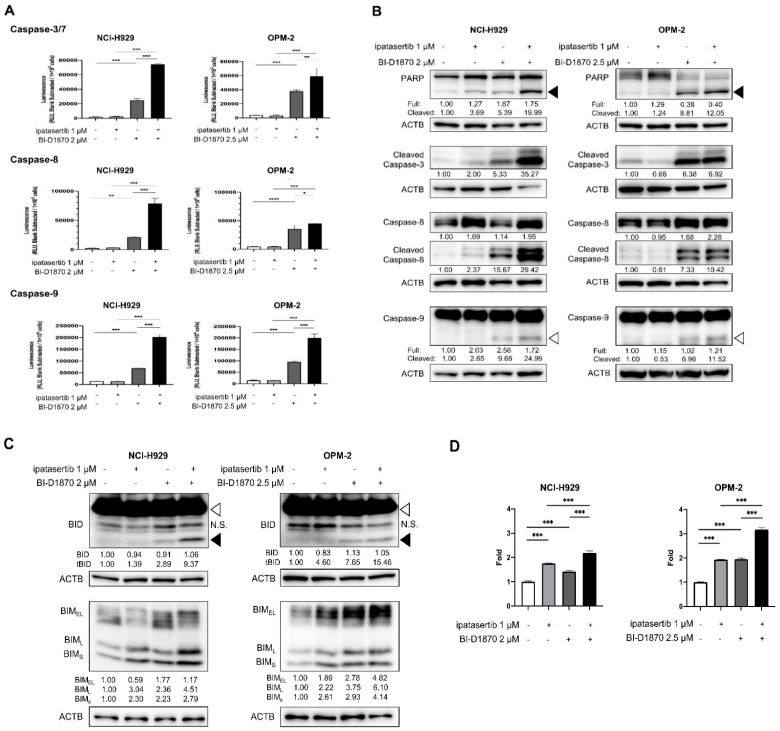
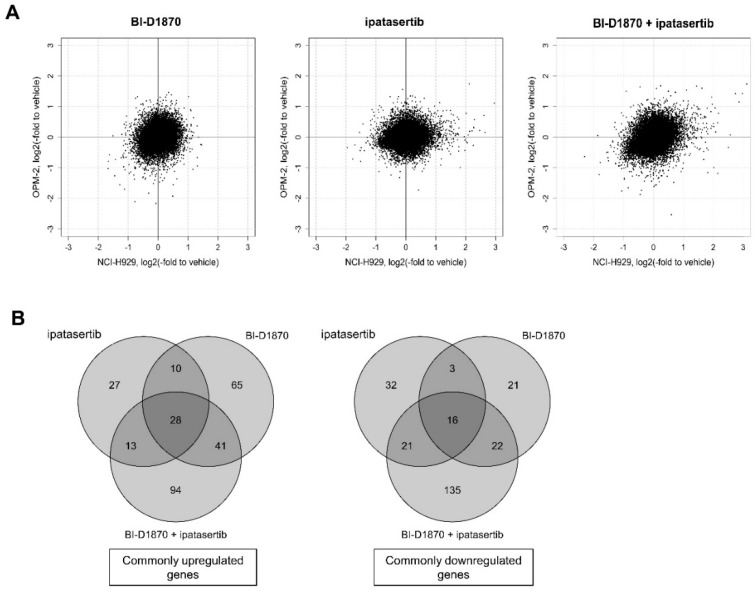
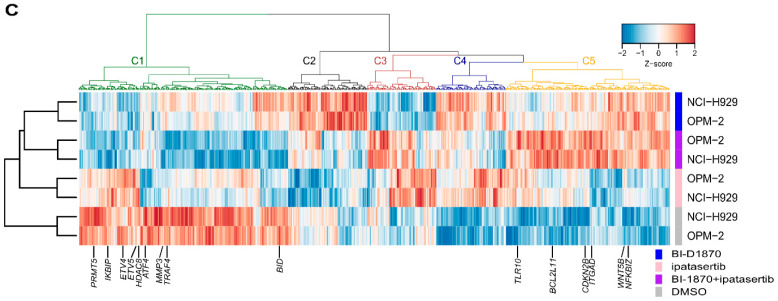
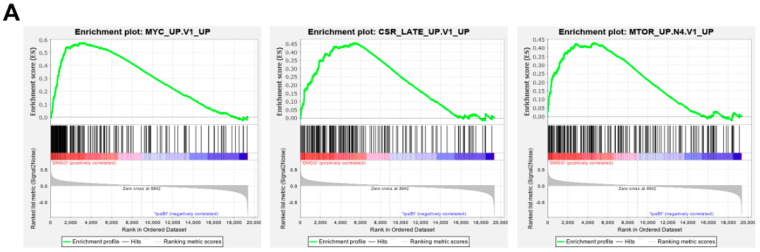
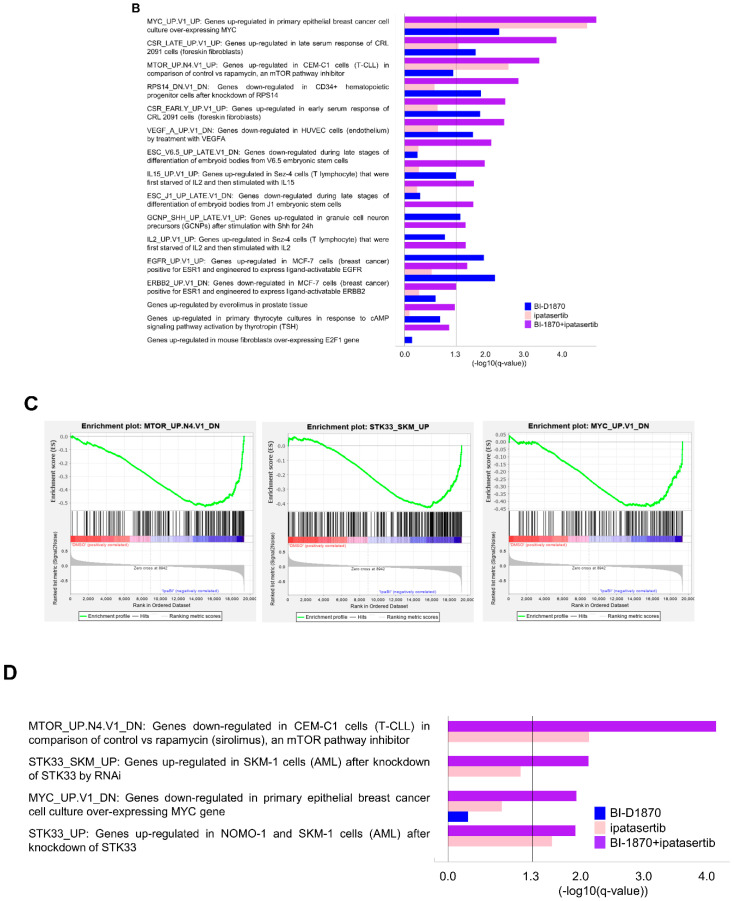
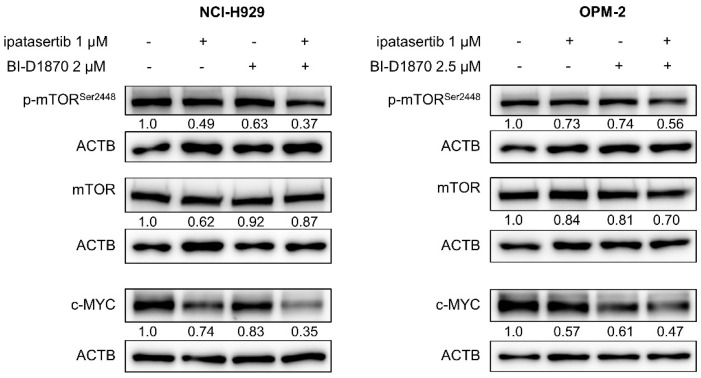
Multiple myeloma (MM) is characterized by remarkable cytogenetic/molecular heterogeneity among patients and intraclonal diversity even in a single patient. We previously demonstrated that PDPK1, the master kinase of series of AGC kinases, is universally active in MM, and plays pivotal roles in cell proliferation and cell survival of myeloma cells regardless of the profiles of cytogenetic and genetic abnormalities. This study investigated the therapeutic efficacy and mechanism of action of dual blockade of two major PDPK1 substrates, RSK2 and AKT, in MM. The combinatory treatment of BI-D1870, an inhibitor for N-terminal kinase domain (NTKD) of RSK2, and ipatasertib, an inhibitor for AKT, showed the additive to synergistic anti-tumor effect on human MM-derived cell lines (HMCLs) with active RSK2-NTKD and AKT, by enhancing apoptotic induction with BIM and BID activation. Moreover, the dual blockade of RSK2 and AKT exerted robust molecular effects on critical gene sets associated with myeloma pathophysiologies, such as those with MYC, mTOR, STK33, ribosomal biogenesis, or cell-extrinsic stimuli of soluble factors, in HMCLs. These results provide the biological and molecular rationales for the dual-targeting strategy for RSK2 and AKT, which may overcome the therapeutic difficulty due to cytogenetic/molecular heterogeneity in MM.
Keywords: AKT; MYC; RSK2; gene set enrichment analysis; mTOR; multiple myeloma.
Conflict of interest statement
J.K. received research funding from Kyowa Kirin, Chugai Pharmaceutical, Ono Pharmaceutical, Sanofi, Eisai, Bristol-Myers Squibb (BMS), Sysmex, Dainippon Sumitomo Pharma, Nippon Shinyaku, Abbvie, Teijin and Otsuka Pharmaceutical, has received honoraria from Janssen Pharmaceutical K.K, Kyowa Kirin, Chugai Pharmaceutical, Ono Pharmaceutical, Sanofi, Eisai, Symbio, BMS, Astellas Pharma, Pfizer, Nippon Shinyaku, Daiichi Sankyo, Dainippon Sumitomo Pharma, Abbvie and Otsuka Pharmaceutical, and is a consultant for Janssen Pharmaceutical K.K, and BMS. T.T. received research funding from Nippon Shinyaku.
Figures
Similar articles
-
Robust anti-myeloma effect of TAS0612, an RSK/AKT/S6K inhibitor, with venetoclax regardless of cytogenetic abnormalities.Leukemia. 2025 Jan;39(1):211-221. doi: 10.1038/s41375-024-02439-9. Epub 2024 Oct 22. Leukemia. 2025. PMID: 39438587
-
Serine-227 in the N-terminal kinase domain of RSK2 is a potential therapeutic target for mantle cell lymphoma.Cancer Med. 2020 Jul;9(14):5185-5199. doi: 10.1002/cam4.3136. Epub 2020 May 18. Cancer Med. 2020. PMID: 32420699 Free PMC article.
-
RSK2(Ser227) at N-terminal kinase domain is a potential therapeutic target for multiple myeloma.Mol Cancer Ther. 2012 Dec;11(12):2600-9. doi: 10.1158/1535-7163.MCT-12-0605. Epub 2012 Sep 25. Mol Cancer Ther. 2012. PMID: 23012246
-
Targeting RSK2 in Cancer Therapy: A Review of Natural Products.Anticancer Agents Med Chem. 2025;25(1):35-41. doi: 10.2174/0118715206329546240830055233. Anticancer Agents Med Chem. 2025. PMID: 39248063 Review.
-
PI3K/AKT/mTOR pathway in multiple myeloma: from basic biology to clinical promise.Leuk Lymphoma. 2018 Nov;59(11):2524-2534. doi: 10.1080/10428194.2017.1421760. Epub 2018 Jan 11. Leuk Lymphoma. 2018. PMID: 29322846 Review.
Cited by
-
Multiple myeloma: signaling pathways and targeted therapy.Mol Biomed. 2024 Jul 4;5(1):25. doi: 10.1186/s43556-024-00188-w. Mol Biomed. 2024. PMID: 38961036 Free PMC article. Review.
-
Predicting the Progression from Asymptomatic to Symptomatic Multiple Myeloma and Stage Classification Using Gene Expression Data.Cancers (Basel). 2025 Jan 20;17(2):332. doi: 10.3390/cancers17020332. Cancers (Basel). 2025. PMID: 39858113 Free PMC article.
-
Post-Translational Modifications in Multiple Myeloma: Mechanisms of Drug Resistance and Therapeutic Opportunities.Biomolecules. 2025 May 12;15(5):702. doi: 10.3390/biom15050702. Biomolecules. 2025. PMID: 40427595 Free PMC article. Review.
-
Triple targeting of RSK, AKT, and S6K as pivotal downstream effectors of PDPK1 by TAS0612 in B-cell lymphomas.Cancer Sci. 2023 Dec;114(12):4691-4705. doi: 10.1111/cas.15995. Epub 2023 Oct 15. Cancer Sci. 2023. PMID: 37840379 Free PMC article.
-
A first-in-human phase I study of TAS-117, an allosteric AKT inhibitor, in patients with advanced solid tumors.Cancer Chemother Pharmacol. 2024 Jun;93(6):605-616. doi: 10.1007/s00280-023-04631-7. Epub 2024 Feb 27. Cancer Chemother Pharmacol. 2024. PMID: 38411735 Free PMC article. Clinical Trial.
References
-
- Fujibayashi Y., Isa R., Nishiyama D., Sakamoto-Inada N., Kawasumi N., Yamaguchi J., Kuwahara-Ota S., Matsumura-Kimoto Y., Tsukamoto T., Chinen Y., et al. Aberrant BUB1 Overexpression Promotes Mitotic Segregation Errors and Chromosomal Instability in Multiple Myeloma. Cancers. 2020;12:2206. doi: 10.3390/cancers12082206. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
