

Ubiquitination of Receptorsomes, Frontline of Plant Immunity
- PMID: 35328358
- PMCID: PMC8948693
- DOI: 10.3390/ijms23062937
Ubiquitination of Receptorsomes, Frontline of Plant Immunity
Abstract
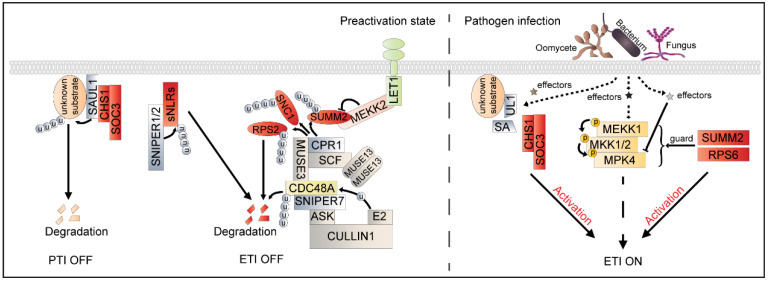
Sessile plants are constantly exposed to myriads of unfavorable invading organisms with different lifestyles. To survive, plants have evolved plasma membrane-resident pattern recognition receptors (PRRs) and intracellular nucleotide-binding domain leucine-rich repeat receptors (NLRs) to initiate sophisticated downstream immune responses. Ubiquitination serves as one of the most important and prevalent posttranslational modifications (PTMs) to fine-tune plant immune responses. Over the last decade, remarkable progress has been made in delineating the critical roles of ubiquitination in plant immunity. In this review, we highlight recent advances in the understanding of ubiquitination in the modulation of plant immunity, with a particular focus on ubiquitination in the regulation of receptorsomes, and discuss how ubiquitination and other PTMs act in concert to ensure rapid, proper, and robust immune responses.
Keywords: plant immunity; posttranslational modifications (PTMs); receptorsomes; ubiquitination.
Conflict of interest statement
The authors declare no conflict of interest. All authors have read and agreed to the published version of the manuscript.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
