

Human C1q Regulates Influenza A Virus Infection and Inflammatory Response via Its Globular Domain
- PMID: 35328462
- PMCID: PMC8949502
- DOI: 10.3390/ijms23063045
Human C1q Regulates Influenza A Virus Infection and Inflammatory Response via Its Globular Domain
Abstract
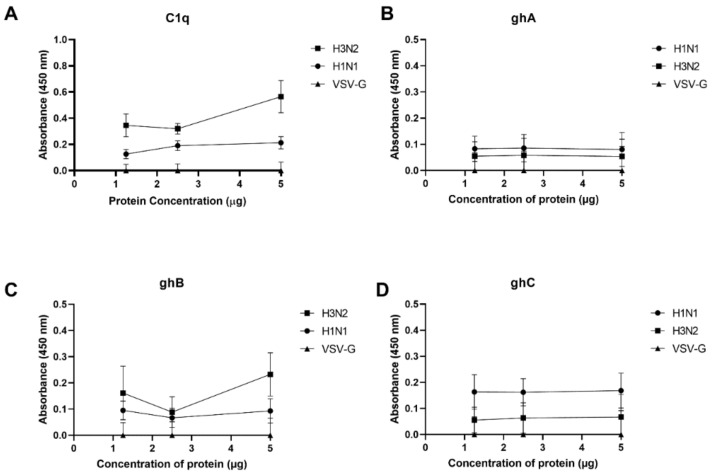
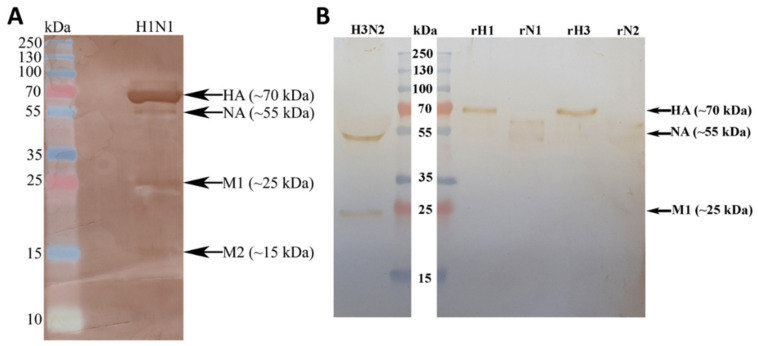
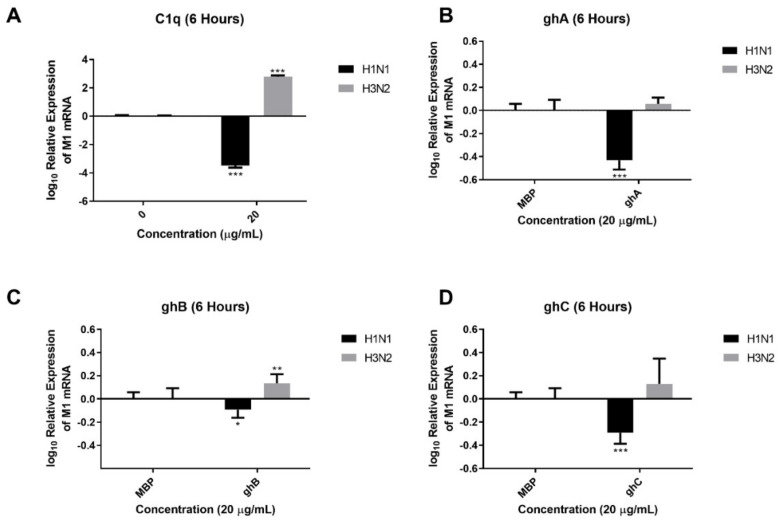
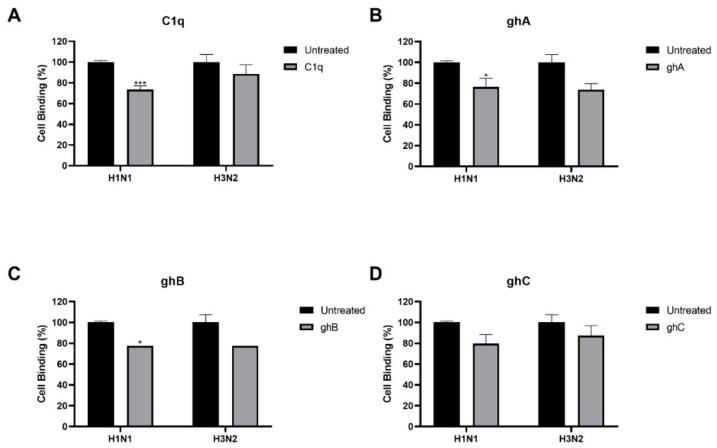
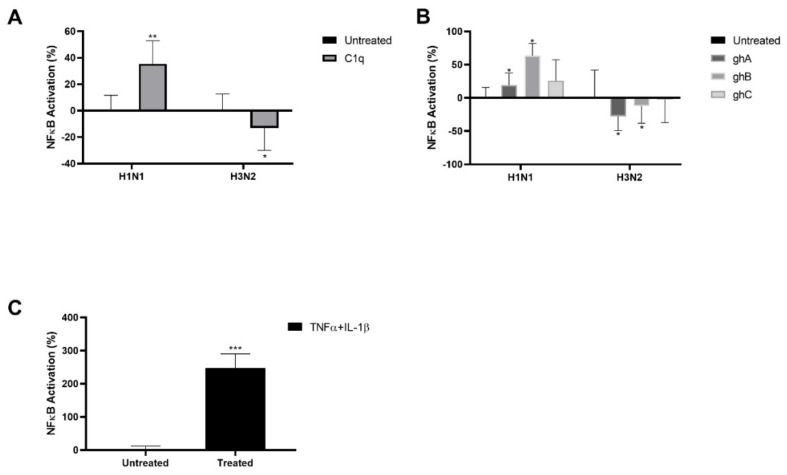
The Influenza A virus (IAV) is a severe respiratory pathogen. C1q is the first subcomponent of the complement system's classical pathway. C1q is composed of 18 polypeptide chains. Each of these chains contains a collagen-like region located at the N terminus, and a C-terminal globular head region organized as a heterotrimeric structure (ghA, ghB and ghC). This study was aimed at investigating the complement activation-independent modulation by C1q and its individual recombinant globular heads against IAV infection. The interaction of C1q and its recombinant globular heads with IAV and its purified glycoproteins was examined using direct ELISA and far-Western blotting analysis. The effect of the complement proteins on IAV replication kinetics and immune modulation was assessed by qPCR. The IAV entry inhibitory properties of C1q and its recombinant globular heads were confirmed using cell binding and luciferase reporter assays. C1q bound IAV virions via HA, NA and M1 IAV proteins, and suppressed replication in H1N1, while promoting replication in H3N2-infected A549 cells. C1q treatment further triggered an anti-inflammatory response in H1N1 and pro-inflammatory response in H3N2-infected cells as evident from differential expression of TNF-α, NF-κB, IFN-α, IFN-β, IL-6, IL-12 and RANTES. Furthermore, C1q treatment was found to reduce luciferase reporter activity of MDCK cells transfected with H1N1 pseudotyped lentiviral particles, indicative of an entry inhibitory role of C1q against infectivity of IAV. These data appear to demonstrate the complement-independent subtype specific modulation of IAV infection by locally produced C1q.
Keywords: RNA viruses; classical pathway; complement; cytokine storm; human C1q; immune evasion; influenza A virus; innate immunity.
Conflict of interest statement
The authors report no declarations of interest.
Figures

References
-
- Simonsen L., Viboud C., Taylor R.J., Miller M.A. The Epidemiology of Influenza and Its Control. In: Rappuoli R., Del Giudice G., editors. Influenza Vaccines for the Future. Springer; Basel, Switzerland: 2011. pp. 27–54.
-
- Bresee J.S., Fry A.M., Sambhara S., Cox N.J. Inactivated Influenza Vaccines. In: Plotkin S.A., Orenstein W.A., Offit P.A., Edwards K.M., editors. Plotkin’s Vaccines. 7th ed. Elsevier; Amsterdam, The Netherlands: 2018. pp. 456–488.e21.
-
- García-Sastre A. Influenza. In: Schaechter M., editor. Encyclopedia of Microbiology. 3rd ed. Academic Press; Oxford, UK: 2009. pp. 414–420.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
