

Sunflower WRINKLED1 Plays a Key Role in Transcriptional Regulation of Oil Biosynthesis
- PMID: 35328473
- PMCID: PMC8951541
- DOI: 10.3390/ijms23063054
Sunflower WRINKLED1 Plays a Key Role in Transcriptional Regulation of Oil Biosynthesis
Abstract
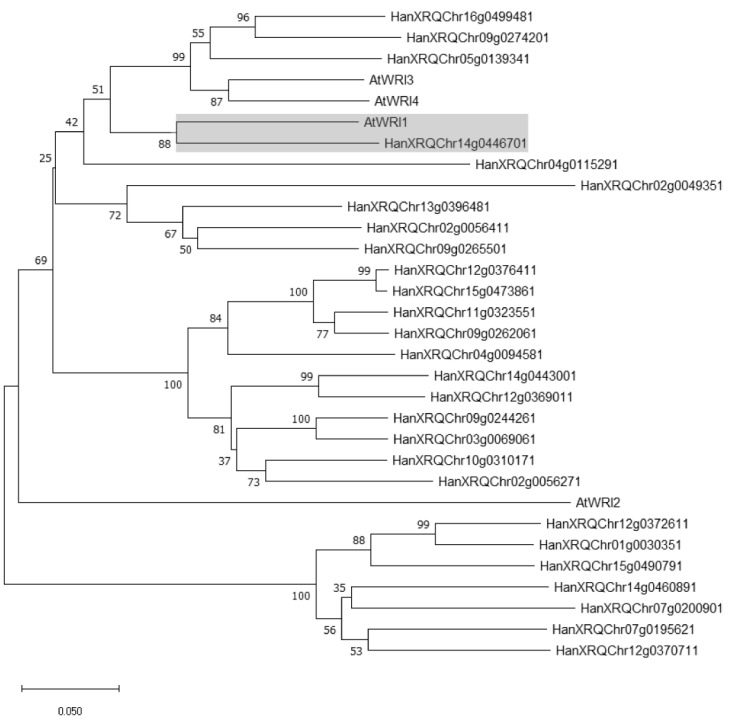
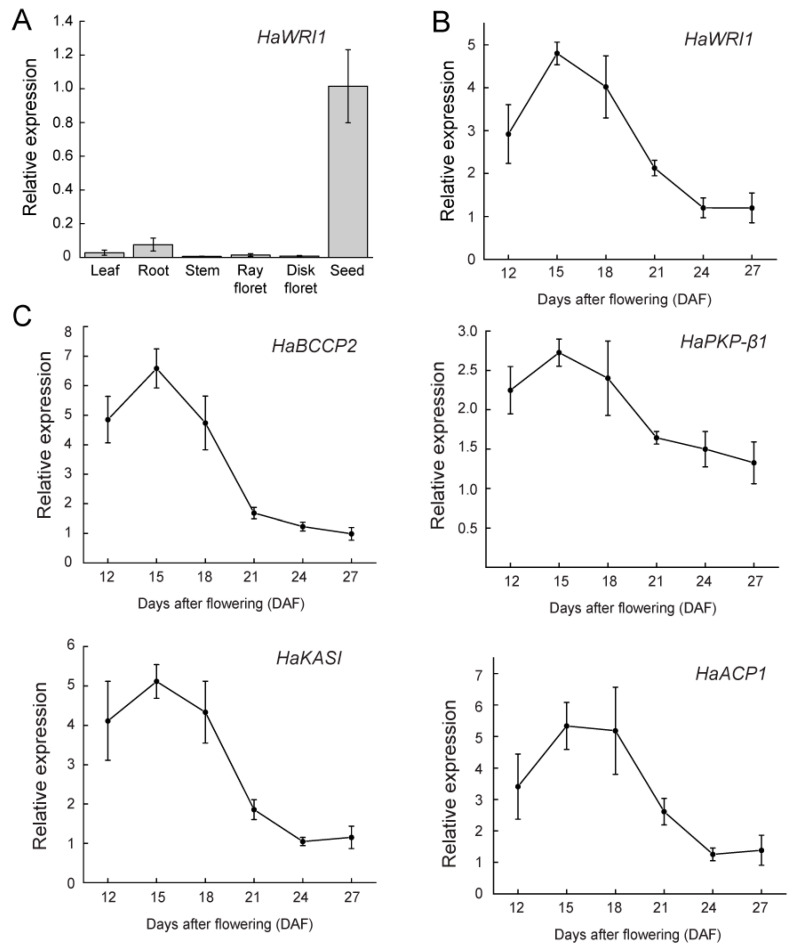
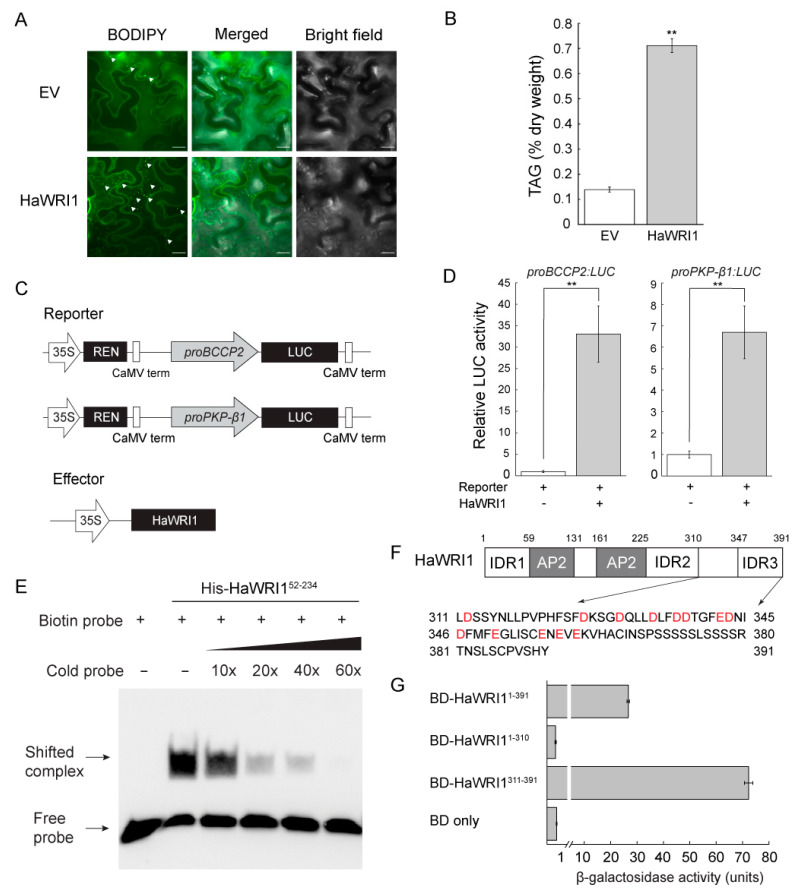
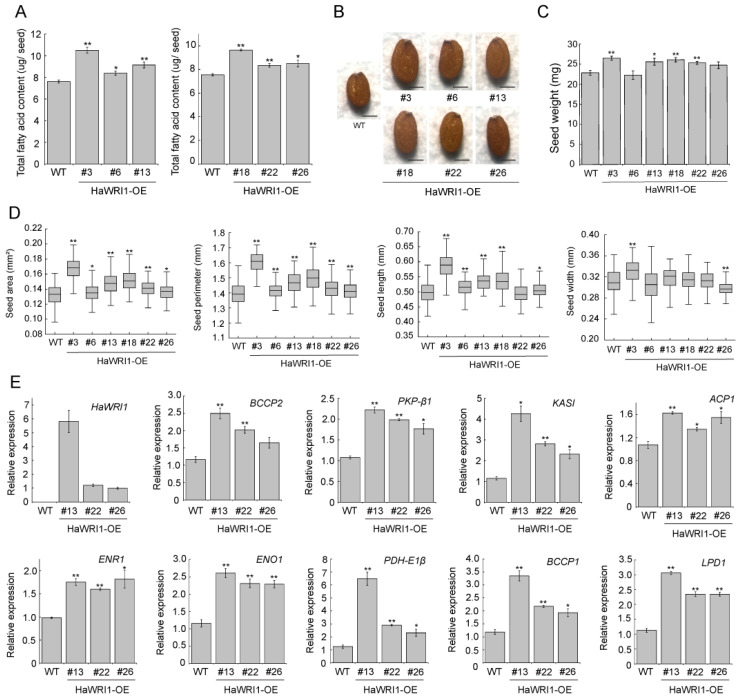
Sunflower (Helianthus annuus) is one of the most important oilseed crops worldwide. However, the transcriptional regulation underlying oil accumulation in sunflower is not fully understood. WRINKLED1 (WRI1) is an essential transcription factor governing oil accumulation in plant cells. Here, we identify and characterize a sunflower ortholog of WRI1 (HaWRI1), which is highly expressed in developing seeds. Transient production of HaWRI1 stimulated substantial oil accumulation in Nicotiana benthamiana leaves. Dual-luciferase reporter assay, electrophoretic mobility shift assay, fatty acid quantification, and gene expression analysis demonstrate that HaWRI1 acts as a pivotal transcription factor controlling the expression of genes involved in late glycolysis and fatty acid biosynthesis. HaWRI1 directly binds to the cis-element, AW-box, in the promoter of biotin carboxyl carrier protein isoform 2 (BCCP2). In addition, we characterize an 80 amino-acid C-terminal domain of HaWRI1 that is crucial for transactivation. Moreover, seed-specific overexpression of HaWRI1 in Arabidopsis plants leads to enhanced seed oil content as well as upregulation of the genes involved in fatty acid biosynthesis. Taken together, our work demonstrates that HaWRI1 plays a pivotal role in the transcriptional control of seed oil accumulation, providing a potential target for bioengineering sunflower oil yield improvement.
Keywords: WRI1; gene expression; plant oil biosynthesis; sunflower; transactivation; transcription factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Role of WRINKLED1 in the transcriptional regulation of glycolytic and fatty acid biosynthetic genes in Arabidopsis.Plant J. 2009 Dec;60(6):933-47. doi: 10.1111/j.1365-313X.2009.04011.x. Plant J. 2009. PMID: 19719479
-
WRINKLED1 Positively Regulates Oil Biosynthesis in Carya cathayensis.J Agric Food Chem. 2023 May 3;71(17):6763-6774. doi: 10.1021/acs.jafc.3c00358. Epub 2023 Apr 4. J Agric Food Chem. 2023. PMID: 37014130
-
Transcription factor bZIP52 modulates Arabidopsis seed oil biosynthesis through interaction with WRINKLED1.Plant Physiol. 2023 Aug 3;192(4):2628-2639. doi: 10.1093/plphys/kiad270. Plant Physiol. 2023. PMID: 37148285
-
Genetic possibilities for altering sunflower oil quality to obtain novel oils.Can J Physiol Pharmacol. 2008 Apr;86(4):215-21. doi: 10.1139/Y08-008. Can J Physiol Pharmacol. 2008. PMID: 18418432 Review.
-
WRINKLED1 transcription factor: How much do we know about its regulatory mechanism?Plant Sci. 2018 Jul;272:153-156. doi: 10.1016/j.plantsci.2018.04.013. Epub 2018 Apr 18. Plant Sci. 2018. PMID: 29807586 Review.
Cited by
-
GmWRI1c Increases Palmitic Acid Content to Regulate Seed Oil Content and Nodulation in Soybean (Glycine max).Int J Mol Sci. 2022 Nov 9;23(22):13793. doi: 10.3390/ijms232213793. Int J Mol Sci. 2022. PMID: 36430287 Free PMC article.
-
Transcriptional regulation of oil biosynthesis in seed plants: Current understanding, applications, and perspectives.Plant Commun. 2022 Sep 12;3(5):100328. doi: 10.1016/j.xplc.2022.100328. Epub 2022 Apr 20. Plant Commun. 2022. PMID: 35605194 Free PMC article. Review.
-
Transcriptome Analysis Reveals Key Genes Involved in Fatty Acid and Triacylglycerol Accumulation in Developing Sunflower Seeds.Genes (Basel). 2025 Mar 29;16(4):393. doi: 10.3390/genes16040393. Genes (Basel). 2025. PMID: 40282355 Free PMC article.
-
Progress in understanding and improving oil content and quality in seeds.Front Plant Sci. 2023 Jan 26;14:1116894. doi: 10.3389/fpls.2023.1116894. eCollection 2023. Front Plant Sci. 2023. PMID: 36778708 Free PMC article. Review.
-
Determination of superior Pistacia chinensis accession with high-quality seed oil and biodiesel production and revelation of LEC1/WRI1-mediated high oil accumulative mechanism for better developing woody biodiesel.BMC Plant Biol. 2023 May 19;23(1):268. doi: 10.1186/s12870-023-04267-y. BMC Plant Biol. 2023. PMID: 37208597 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
