

PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin
- PMID: 35328480
- PMCID: PMC8954402
- DOI: 10.3390/ijms23063059
PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin
Abstract
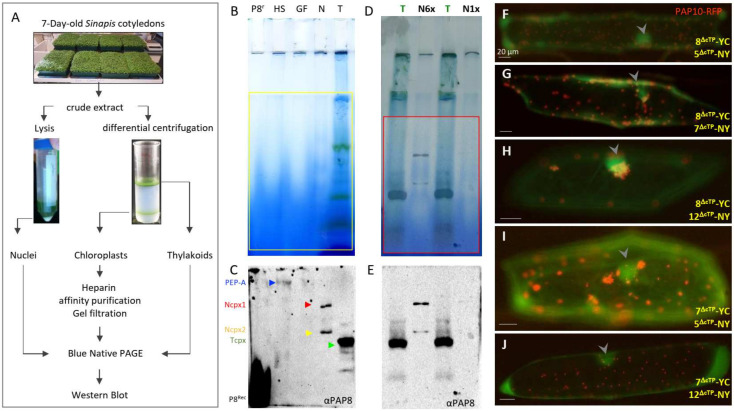
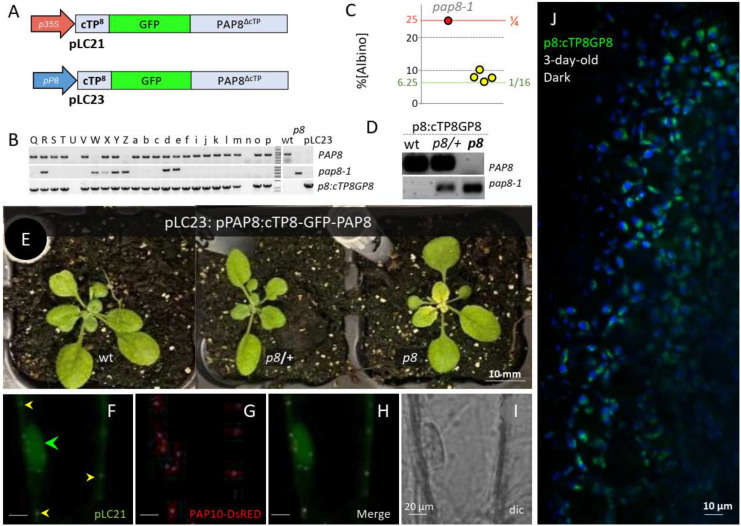
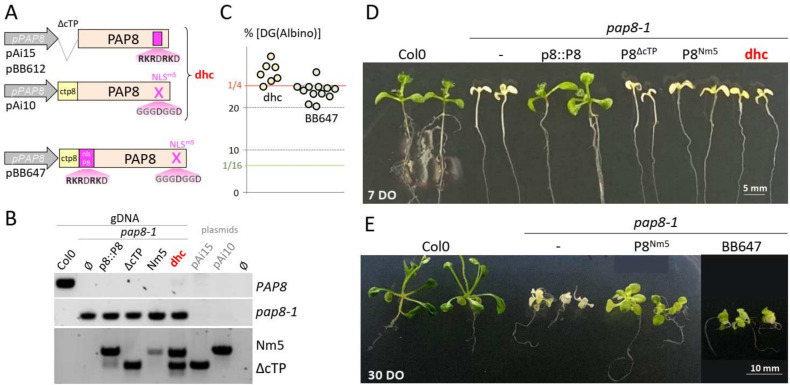
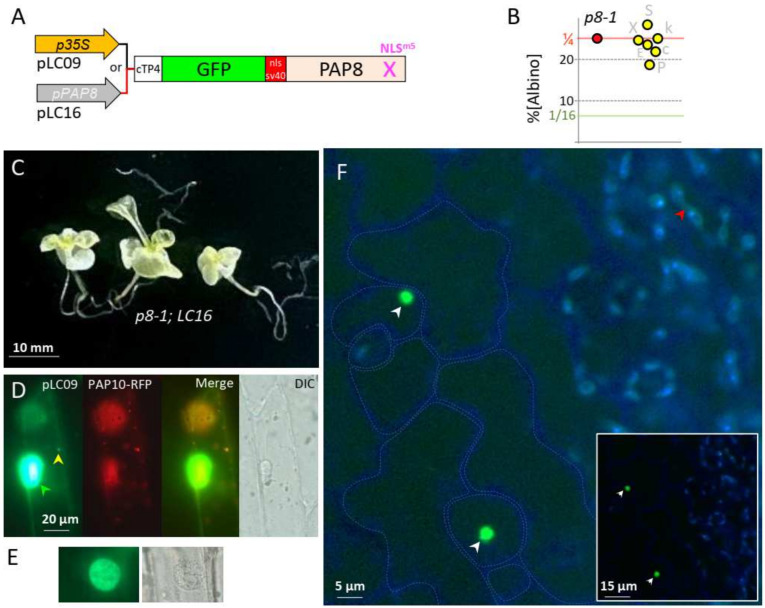
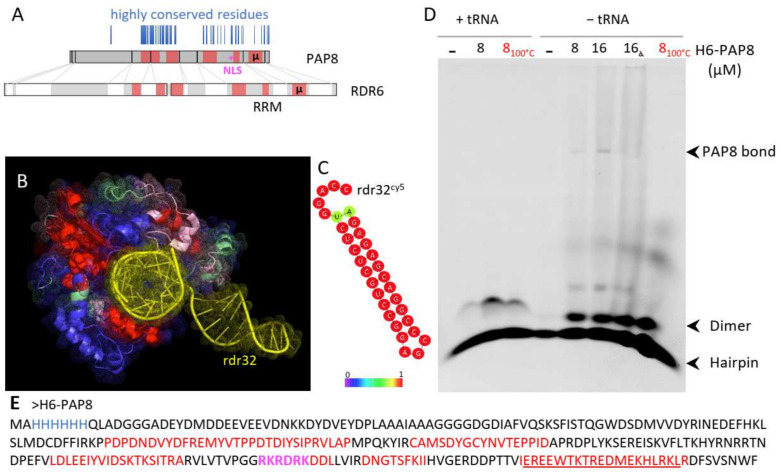
Chloroplast biogenesis depends on a complex transcriptional program involving coordinated expression of plastid and nuclear genes. In particular, photosynthesis-associated plastid genes are expressed by the plastid-encoded polymerase (PEP) that undergoes a structural rearrangement during chloroplast formation. The prokaryotic-type core enzyme is rebuilt into a larger complex by the addition of nuclear-encoded PEP-associated proteins (PAP1 to PAP12). Among the PAPs, some have been detected in the nucleus (PAP5 and PAP8), where they could serve a nuclear function required for efficient chloroplast biogenesis. Here, we detected PAP8 in a large nuclear subcomplex that may include other subunits of the plastid-encoded RNA polymerase. We have made use of PAP8 recombinant proteins in Arabidopsis thaliana to decouple its nucleus- and chloroplast-associated functions and found hypomorphic mutants pointing at essential amino acids. While the origin of the PAP8 gene remained elusive, we have found in its sequence a micro-homologous domain located within a large structural homology with a rhinoviral RNA-dependent RNA polymerase, highlighting potential RNA recognition motifs in PAP8. PAP8 in vitro RNA binding activity suggests that this domain is functional. Hence, we propose that the acquisition of PAPs may have occurred during evolution by different routes, including lateral gene transfer.
Keywords: Arabidopsis; PAP8/pTAC6; PEP/PAPs; RNA recognition motif; biogenesis; chloroplast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Nucleo-plastidic PAP8/pTAC6 couples chloroplast formation with photomorphogenesis.EMBO J. 2020 Nov 16;39(22):e104941. doi: 10.15252/embj.2020104941. Epub 2020 Oct 1. EMBO J. 2020. PMID: 33001465 Free PMC article.
-
PAP genes are tissue- and cell-specific markers of chloroplast development.Planta. 2018 Sep;248(3):629-646. doi: 10.1007/s00425-018-2924-8. Epub 2018 May 31. Planta. 2018. PMID: 29855700
-
PAP1 and PAP7 are required for association of plastid-encoded RNA polymerase with DNA.Plant Mol Biol. 2024 Sep 20;114(5):100. doi: 10.1007/s11103-024-01498-x. Plant Mol Biol. 2024. PMID: 39302509
-
The plastid transcription machinery and its coordination with the expression of nuclear genome: Plastid-Encoded Polymerase, Nuclear-Encoded Polymerase and the Genomes Uncoupled 1-mediated retrograde communication.Philos Trans R Soc Lond B Biol Sci. 2020 Jun 22;375(1801):20190399. doi: 10.1098/rstb.2019.0399. Epub 2020 May 4. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32362266 Free PMC article. Review.
-
Chloroplast RNA polymerases: Role in chloroplast biogenesis.Biochim Biophys Acta. 2015 Sep;1847(9):761-9. doi: 10.1016/j.bbabio.2015.02.004. Epub 2015 Feb 11. Biochim Biophys Acta. 2015. PMID: 25680513 Review.
Cited by
-
Chloroplast gene expression: Recent advances and perspectives.Plant Commun. 2023 Sep 11;4(5):100611. doi: 10.1016/j.xplc.2023.100611. Epub 2023 May 4. Plant Commun. 2023. PMID: 37147800 Free PMC article. Review.
-
Three-Dimensional Envelope and Subunit Interactions of the Plastid-Encoded RNA Polymerase from Sinapis alba.Int J Mol Sci. 2022 Aug 31;23(17):9922. doi: 10.3390/ijms23179922. Int J Mol Sci. 2022. PMID: 36077319 Free PMC article.
-
Light-regulated dual-targeting of NUCLEAR CONTROL OF PEP ACTIVITY establishes photomorphogenesis via interorganellar communication.Plant Physiol. 2025 Sep 1;199(1):kiaf289. doi: 10.1093/plphys/kiaf289. Plant Physiol. 2025. PMID: 40587430 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
