

Dysfunctional cGMP Signaling Leads to Age-Related Retinal Vascular Alterations and Astrocyte Remodeling in Mice
- PMID: 35328488
- PMCID: PMC8954518
- DOI: 10.3390/ijms23063066
Dysfunctional cGMP Signaling Leads to Age-Related Retinal Vascular Alterations and Astrocyte Remodeling in Mice
Abstract
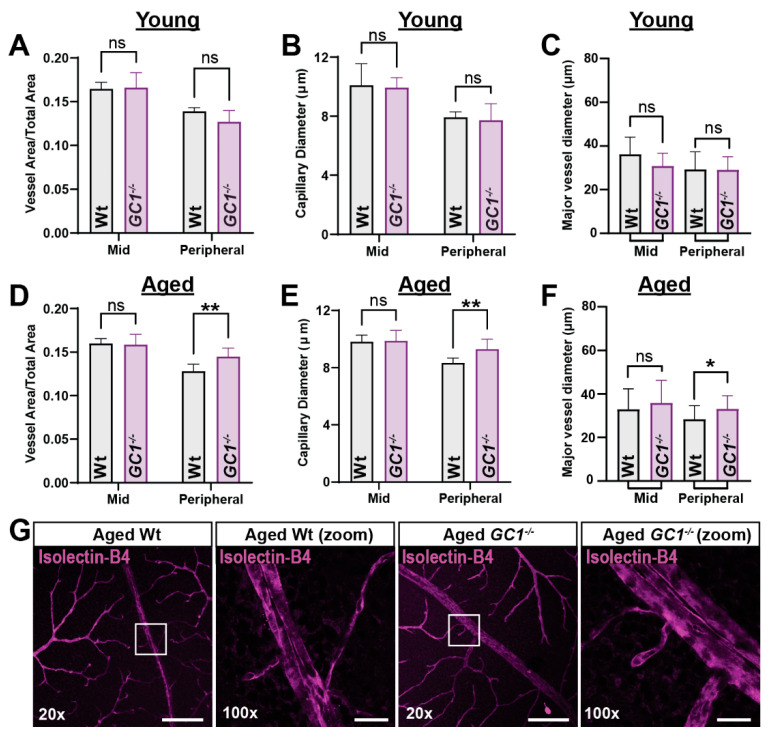
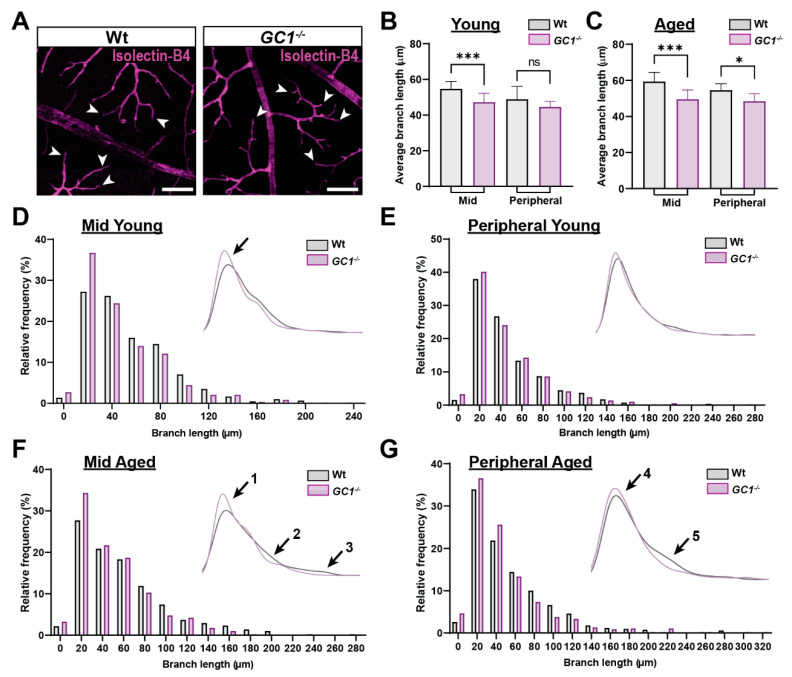
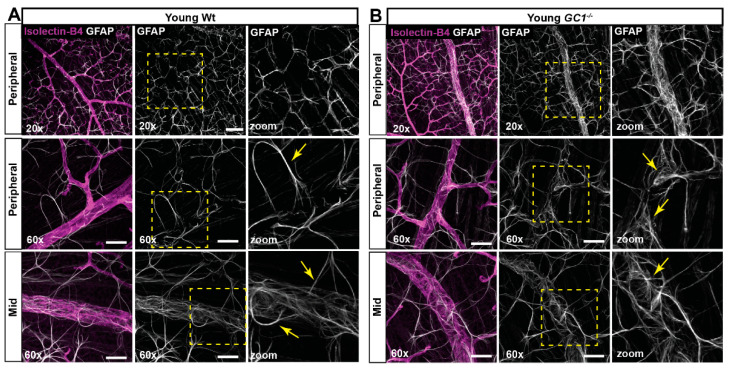
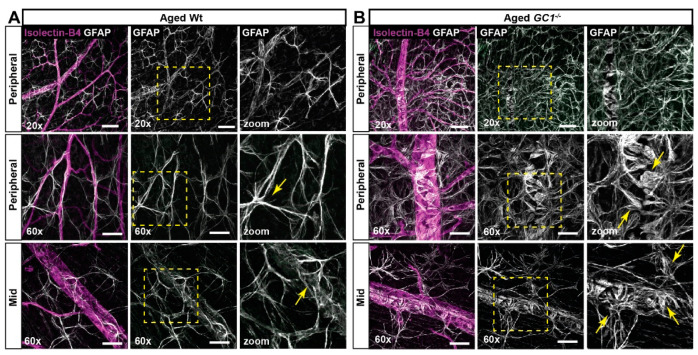
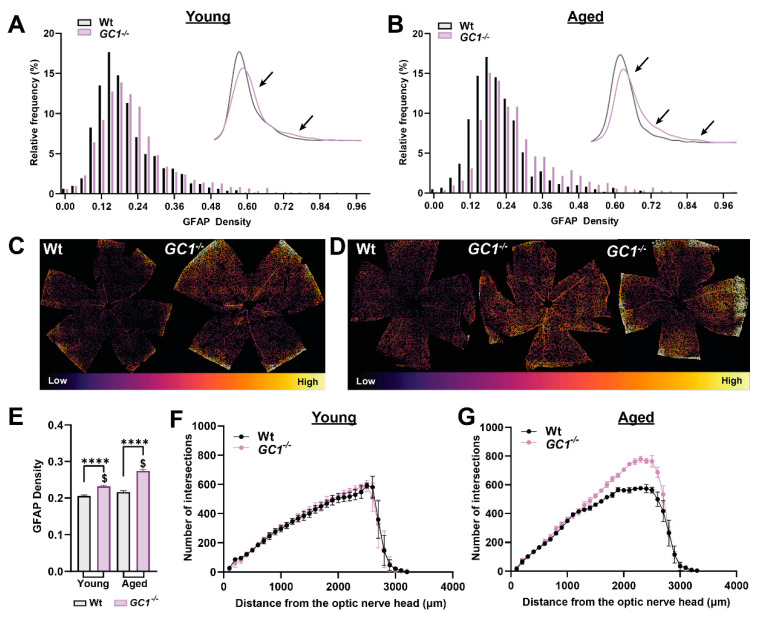
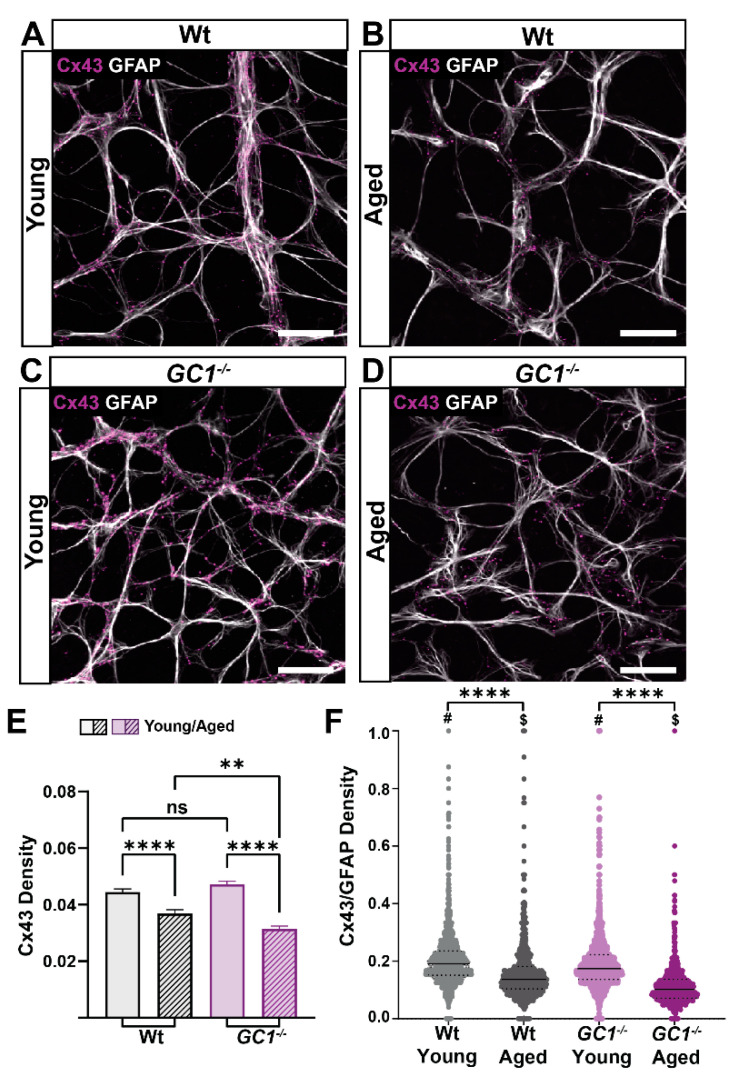
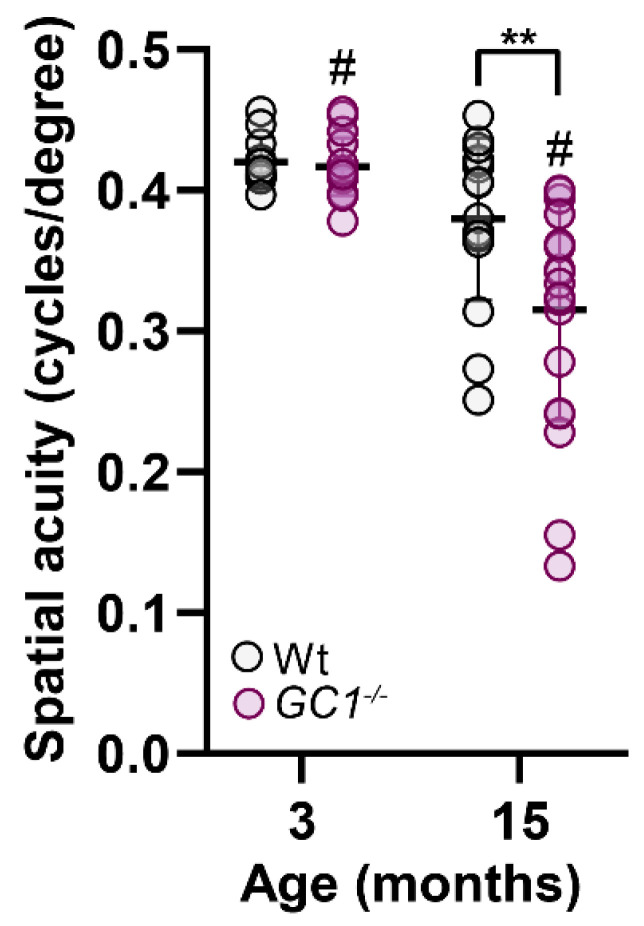
The nitric oxide-guanylyl cyclase-1-cyclic guanylate monophosphate (NO-GC-1-cGMP) pathway is integral to the control of vascular tone and morphology. Mice lacking the alpha catalytic domain of guanylate cyclase (GC1-/-) develop retinal ganglion cell (RGC) degeneration with age, with only modest fluctuations in intraocular pressure (IOP). Increasing the bioavailability of cGMP in GC1-/- mice prevents neurodegeneration independently of IOP, suggesting alternative mechanisms of retinal neurodegeneration. In continuation to these studies, we explored the hypothesis that dysfunctional cGMP signaling leads to changes in the neurovascular unit that may contribute to RGC degeneration. We assessed retinal vasculature and astrocyte morphology in young and aged GC1-/- and wild type mice. GC1-/- mice exhibit increased peripheral retinal vessel dilation and shorter retinal vessel branching with increasing age compared to Wt mice. Astrocyte cell morphology is aberrant, and glial fibrillary acidic protein (GFAP) density is increased in young and aged GC1-/- mice, with areas of dense astrocyte matting around blood vessels. Our results suggest that proper cGMP signaling is essential to retinal vessel morphology with increasing age. Vascular changed are preceded by alterations in astrocyte morphology which may together contribute to retinal neurodegeneration and loss of visual acuity observed in GC1-/- mice.
Keywords: astrocyte; blood retinal barrier; connexin; gap junctions; neurodegeneration; neurovascular unit; retinal ganglion cell; retinal vasculature.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kemp-Harper B., Schmidt H.H. cGMP: Generators, Effectors and Therapeutic Implications. Springer; Berlin/Heidelberg, Germany: 2009. cGMP in the Vasculature; pp. 447–467. Handbook of Experimental Pharmacology 191.
-
- Kang J.H., Wiggs J.L., Rosner B.A., Hankinson S.E., Abdrabou W., Fan B.J., Haines J., Pasquale L.R. Endothelial nitric oxide synthase gene variants and primary open-angle glaucoma: Interactions with sex and postmenopausal hormone use. Investig. Ophthalmol. Vis. Sci. 2010;51:971–979. doi: 10.1167/iovs.09-4266. - DOI - PMC - PubMed
-
- Magalhaes da Silva T., Rocha A.V., Lacchini R., Marques C.R., Silva E.S., Tanus-Santos J.E., Rios-Santos F. Association of polymorphisms of endothelial nitric oxide synthase (eNOS) gene with the risk of primary open angle glaucoma in a Brazilian population. Gene. 2012;502:142–146. doi: 10.1016/j.gene.2012.04.047. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
