

Mono a Mano: ZBP1's Love-Hate Relationship with the Kissing Virus
- PMID: 35328502
- PMCID: PMC8955656
- DOI: 10.3390/ijms23063079
Mono a Mano: ZBP1's Love-Hate Relationship with the Kissing Virus
Abstract
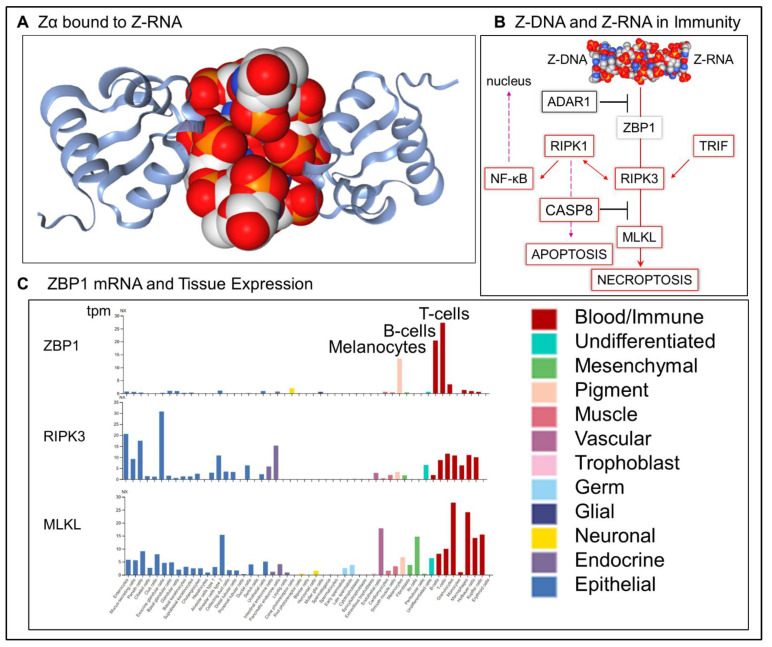
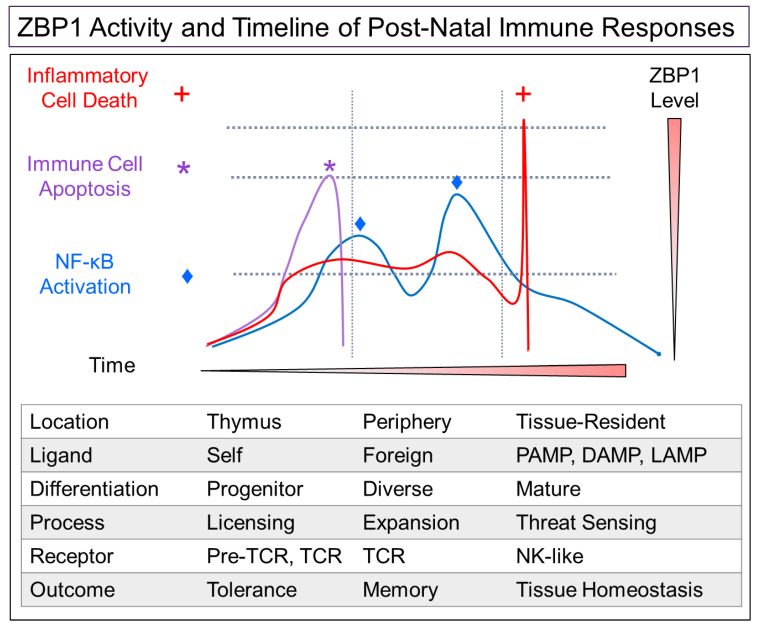
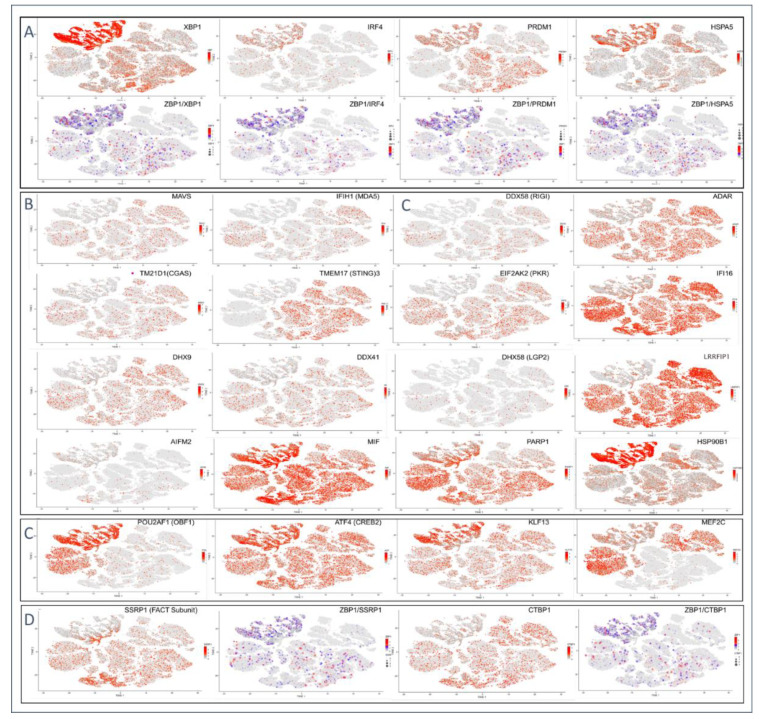
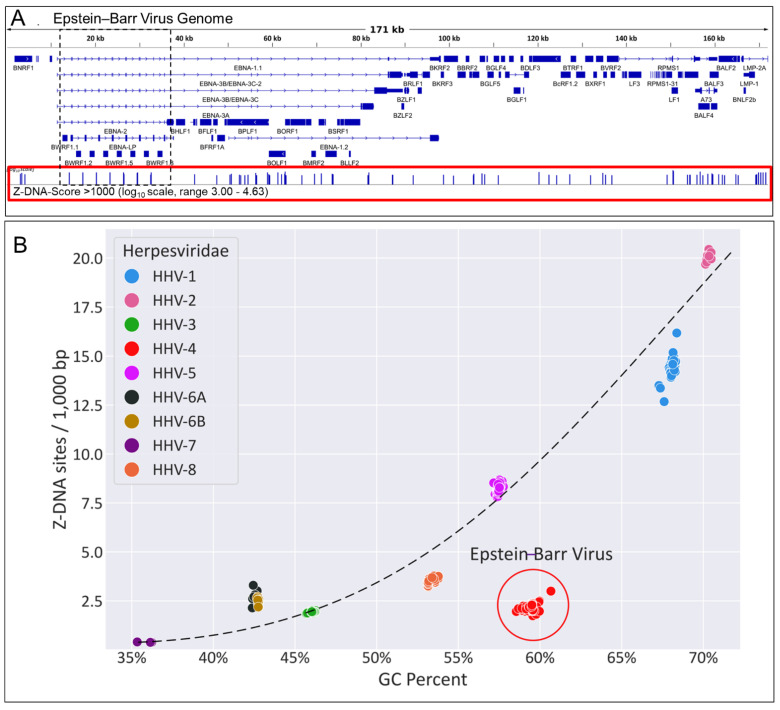
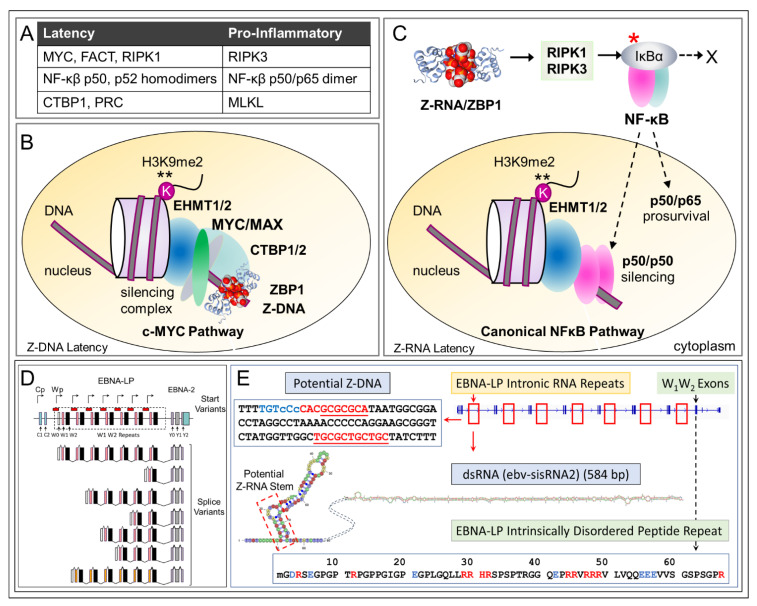
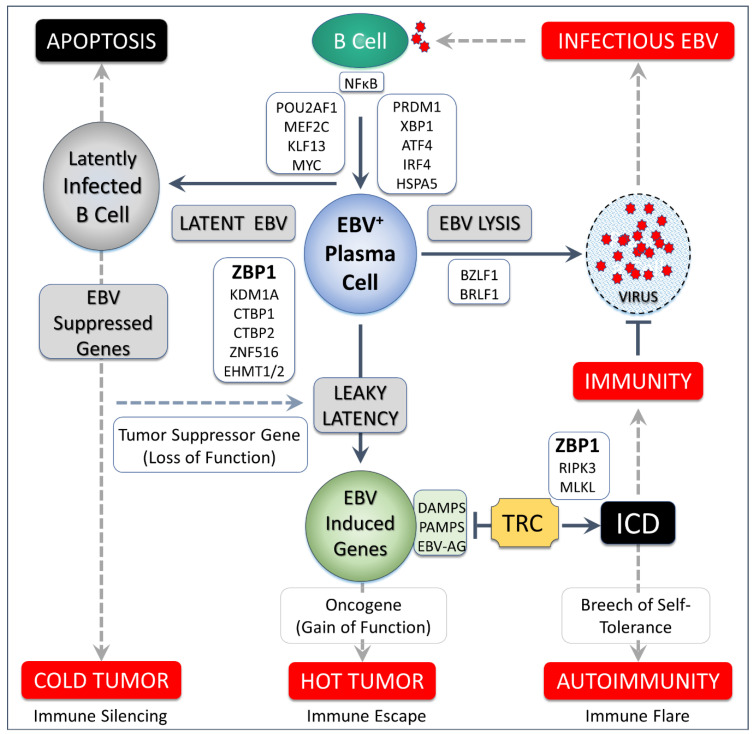
Z-DNA binding protein (ZBP1) very much represents the nuclear option. By initiating inflammatory cell death (ICD), ZBP1 activates host defenses to destroy infectious threats. ZBP1 is also able to induce noninflammatory regulated cell death via apoptosis (RCD). ZBP1 senses the presence of left-handed Z-DNA and Z-RNA (ZNA), including that formed by expression of endogenous retroelements. Viruses such as the Epstein-Barr "kissing virus" inhibit ICD, RCD and other cell death signaling pathways to produce persistent infection. EBV undergoes lytic replication in plasma cells, which maintain detectable levels of basal ZBP1 expression, leading us to suggest a new role for ZBP1 in maintaining EBV latency, one of benefit for both host and virus. We provide an overview of the pathways that are involved in establishing latent infection, including those regulated by MYC and NF-κB. We describe and provide a synthesis of the evidence supporting a role for ZNA in these pathways, highlighting the positive and negative selection of ZNA forming sequences in the EBV genome that underscores the coadaptation of host and virus. Instead of a fight to the death, a state of détente now exists where persistent infection by the virus is tolerated by the host, while disease outcomes such as death, autoimmunity and cancer are minimized. Based on these new insights, we propose actionable therapeutic approaches to unhost EBV.
Keywords: ADAR1; Epstein–Barr virus; Z-DNA; Z-RNA; ZBP1; autoimmune disease; cancer; exhausted T cells; flipons; lymphoma; systemic lupus erythematous.
Conflict of interest statement
Author A.H. is the founder of InsideOutBio, a company that works in the field of immuno-oncology. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Histone Loaders CAF1 and HIRA Restrict Epstein-Barr Virus B-Cell Lytic Reactivation.mBio. 2020 Oct 27;11(5):e01063-20. doi: 10.1128/mBio.01063-20. mBio. 2020. PMID: 33109754 Free PMC article.
-
Regulation and dysregulation of Epstein-Barr virus latency: implications for the development of autoimmune diseases.Autoimmunity. 2008 May;41(4):298-328. doi: 10.1080/08916930802024772. Autoimmunity. 2008. PMID: 18432410 Review.
-
Epstein-Barr virus detection in non-Hodgkin's lymphoma of the oral cavity: an immunocytochemical and in situ hybridization study.Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2001 Aug;92(2):184-93. doi: 10.1067/moe.2001.116155. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2001. PMID: 11505266
-
Interferon-γ-inducible protein 16 (IFI16) is required for the maintenance of Epstein-Barr virus latency.Virol J. 2017 Nov 13;14(1):221. doi: 10.1186/s12985-017-0891-5. Virol J. 2017. PMID: 29132393 Free PMC article.
-
Latent and lytic Epstein-Barr virus replication strategies.Rev Med Virol. 2005 Jan-Feb;15(1):3-15. doi: 10.1002/rmv.441. Rev Med Virol. 2005. PMID: 15386591 Review.
Cited by
-
Bioinformatics-based screening and validation of PANoptosis-related biomarkers in periodontitis.Front Cell Dev Biol. 2025 Jun 19;13:1619002. doi: 10.3389/fcell.2025.1619002. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40612110 Free PMC article.
-
Flipons and small RNAs accentuate the asymmetries of pervasive transcription by the reset and sequence-specific microcoding of promoter conformation.J Biol Chem. 2023 Sep;299(9):105140. doi: 10.1016/j.jbc.2023.105140. Epub 2023 Aug 5. J Biol Chem. 2023. PMID: 37544644 Free PMC article. Review.
-
Z-DNA enhances immunotherapy by triggering death of inflammatory cancer-associated fibroblasts.J Immunother Cancer. 2022 Nov;10(11):e005704. doi: 10.1136/jitc-2022-005704. J Immunother Cancer. 2022. PMID: 36450382 Free PMC article.
-
ZBP1: A Powerful Innate Immune Sensor and Double-Edged Sword in Host Immunity.Int J Mol Sci. 2022 Sep 6;23(18):10224. doi: 10.3390/ijms231810224. Int J Mol Sci. 2022. PMID: 36142136 Free PMC article. Review.
-
Z-RNA and the Flipside of the SARS Nsp13 Helicase: Is There a Role for Flipons in Coronavirus-Induced Pathology?Front Immunol. 2022 Jun 17;13:912717. doi: 10.3389/fimmu.2022.912717. eCollection 2022. Front Immunol. 2022. PMID: 35784331 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
