

CRISPR/Cas9-Based Mutagenesis of Histone H3.1 in Spinal Dynorphinergic Neurons Attenuates Thermal Sensitivity in Mice
- PMID: 35328599
- PMCID: PMC8955318
- DOI: 10.3390/ijms23063178
CRISPR/Cas9-Based Mutagenesis of Histone H3.1 in Spinal Dynorphinergic Neurons Attenuates Thermal Sensitivity in Mice
Abstract
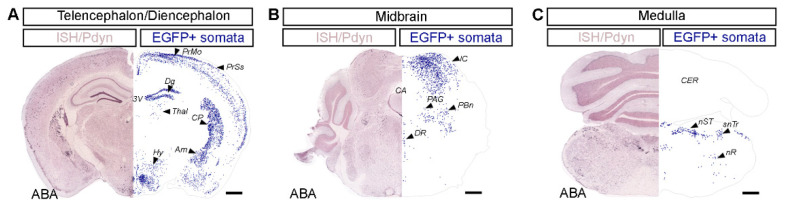
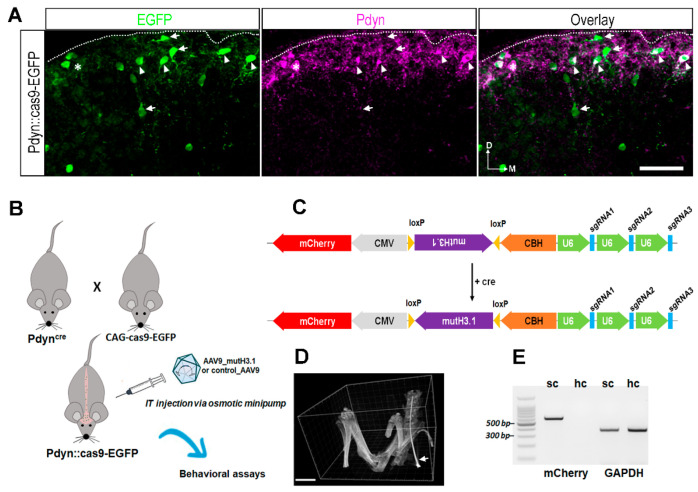
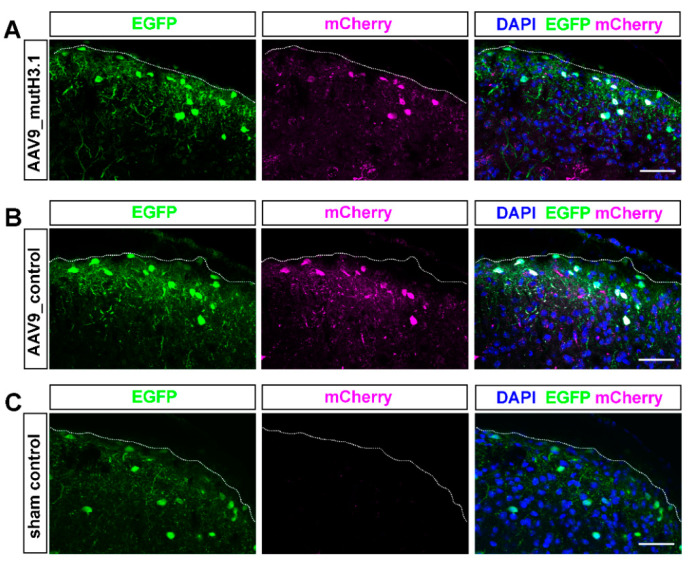
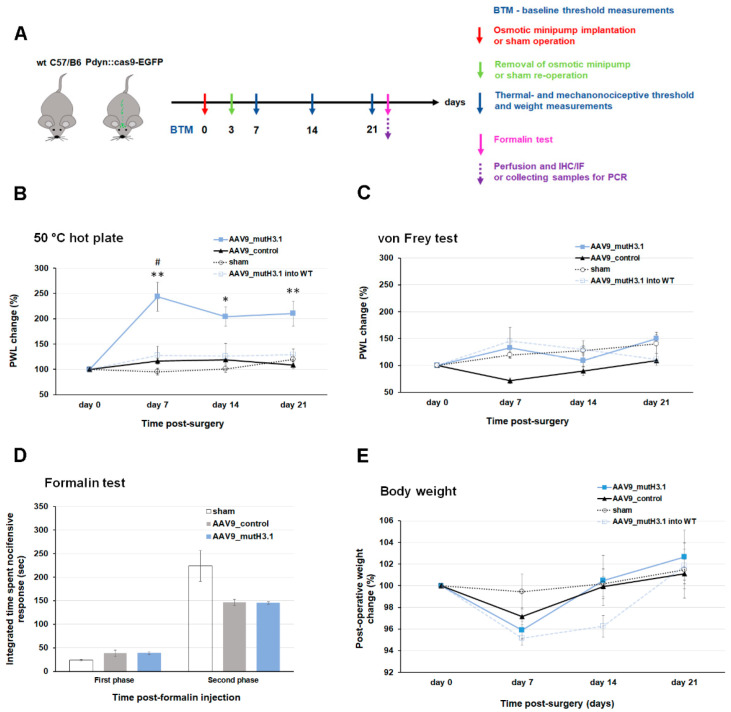
Burn injury is a trauma resulting in tissue degradation and severe pain, which is processed first by neuronal circuits in the spinal dorsal horn. We have recently shown that in mice, excitatory dynorphinergic (Pdyn) neurons play a pivotal role in the response to burn-injury-associated tissue damage via histone H3.1 phosphorylation-dependent signaling. As Pdyn neurons were mostly associated with mechanical allodynia, their involvement in thermonociception had to be further elucidated. Using a custom-made AAV9_mutH3.1 virus combined with the CRISPR/cas9 system, here we provide evidence that blocking histone H3.1 phosphorylation at position serine 10 (S10) in spinal Pdyn neurons significantly increases the thermal nociceptive threshold in mice. In contrast, neither mechanosensation nor acute chemonociception was affected by the transgenic manipulation of histone H3.1. These results suggest that blocking rapid epigenetic tagging of S10H3 in spinal Pdyn neurons alters acute thermosensation and thus explains the involvement of Pdyn cells in the immediate response to burn-injury-associated tissue damage.
Keywords: dynorphinergic neuron; epigenetic regulation; heat sensation; histone; pain; spinal cord.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures
Similar articles
-
Epigenetic Regulation and Molecular Mechanisms of Burn Injury-Induced Nociception in the Spinal Cord of Mice.Int J Mol Sci. 2024 Aug 4;25(15):8510. doi: 10.3390/ijms25158510. Int J Mol Sci. 2024. PMID: 39126078 Free PMC article.
-
Spinal Excitatory Dynorphinergic Interneurons Contribute to Burn Injury-Induced Nociception Mediated by Phosphorylated Histone 3 at Serine 10 in Rodents.Int J Mol Sci. 2021 Feb 25;22(5):2297. doi: 10.3390/ijms22052297. Int J Mol Sci. 2021. PMID: 33669046 Free PMC article.
-
Phosphorylated Histone 3 at Serine 10 Identifies Activated Spinal Neurons and Contributes to the Development of Tissue Injury-Associated Pain.Sci Rep. 2017 Jan 25;7:41221. doi: 10.1038/srep41221. Sci Rep. 2017. PMID: 28120884 Free PMC article.
-
Identification of brain-to-spinal circuits controlling the laterality and duration of mechanical allodynia in mice.Cell Rep. 2023 Apr 25;42(4):112300. doi: 10.1016/j.celrep.2023.112300. Epub 2023 Mar 22. Cell Rep. 2023. PMID: 36952340
-
The NAv1.7 blocker protoxin II reduces burn injury-induced spinal nociceptive processing.J Mol Med (Berl). 2018 Jan;96(1):75-84. doi: 10.1007/s00109-017-1599-0. Epub 2017 Oct 23. J Mol Med (Berl). 2018. PMID: 29063143 Free PMC article.
Cited by
-
Epigenetic Regulation and Molecular Mechanisms of Burn Injury-Induced Nociception in the Spinal Cord of Mice.Int J Mol Sci. 2024 Aug 4;25(15):8510. doi: 10.3390/ijms25158510. Int J Mol Sci. 2024. PMID: 39126078 Free PMC article.
-
Pain modulation in the spinal cord.Front Pain Res (Lausanne). 2022 Sep 13;3:984042. doi: 10.3389/fpain.2022.984042. eCollection 2022. Front Pain Res (Lausanne). 2022. PMID: 36176710 Free PMC article. Review.
-
Roles of Histone H2B, H3 and H4 Variants in Cancer Development and Prognosis.Int J Mol Sci. 2024 Sep 7;25(17):9699. doi: 10.3390/ijms25179699. Int J Mol Sci. 2024. PMID: 39273649 Free PMC article. Review.
-
The Burning Pain Transcriptome in the Mouse Primary Somatosensory Cortex.Int J Mol Sci. 2025 Apr 9;26(8):3538. doi: 10.3390/ijms26083538. Int J Mol Sci. 2025. PMID: 40332032 Free PMC article.
-
Pathology of pain and its implications for therapeutic interventions.Signal Transduct Target Ther. 2024 Jun 8;9(1):155. doi: 10.1038/s41392-024-01845-w. Signal Transduct Target Ther. 2024. PMID: 38851750 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
